Експеримент Мезельсона-Сталя
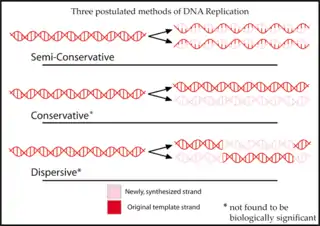
Експеримент Мезельсона-Сталя — експеримент, поставлений Метью Мезельсоном і Франкліном Сталем, щоб довести, що реплікація ДНК є напівконсервативною, тобто коли реплікується подвійна спіраль ДНК, кожна з нових спіралей складається із одного ланцюжка оригінальної спіралі та одного нового ланцюжка.Докази напівконсервативності реплікації ДНК Хромосома- це ниткоподібні щільні тільця, видимі у світловий мікроскоп тільки впродовж поділу клітини. Вони утворюються в результаті ущільнення і спіралізації хроматину. Довжина хромосом залежить від кількості ДНК і білків, а також від ступеня скручування хроматину. На різних ділянках однієї і тієї ж хромосоми спіралізація, компактність її основних елементів неоднакові, з цим пов'язана різна інтенсивність забарвлення окремих ділянок хромосоми. Ділянки, що інтенсивно сприймають барвники, одержали назву гетерохроматичних (утворених із гетерохроматину), вони навіть у період між поділом клітин залишаються компактними, видимими у світловий мікроскоп. Ділянки, що слабко забарвлюються, деконденсуються між поділами клітин, одержали назву еухроматичних (утворених із еухроматину). Встановлено, що еухроматин містить у собі активні гени, а гетерохроматин виконує переважно структурну функцію. Він знаходиться в інтенсивно спіралізованому стані. Гетерохроматин займає однакові ділянки в гомологічних хромосомах: утворює ділянки, що прилягають до центромери і такі, що знаходяться на кінцях хромосом. Втрата ділянок гетерохроматину або мутація може не відбиватися на життєдіяльності клітини. Розрізняють також факультативний гетерохроматин. Він виникає при спіралізації та інактивації однієї з двох гомологічних Х-хромосом. Таким чином утворюється тільце Барра (статевий хроматин). Його утворює одна з двох Х-хро- мосом у жіночих особин ссавців і людини. Кожна молекула ДНК, що утворює хромосому, крім кодуючих послідовностей, повинна містити центромеру, два теломери і точки початку реплікації. Теломерні послідовності запобігають укороченню хромосом, що без них відбувалося б при кожному циклі реплікації ДНК. Центромери служать для вибудовування молекул ДНК на мітотичному веретені під час мітозу. Точки початку реплікації потрібні для формування реплікаційних вилок у S-фазі. Структура метафазної хромосоми. Всі хромосоми під час метафази складаються із двох хроматид, що утворені з максимально спіралізованого хроматину (рис. 1.40). Кожна хроматида - це зв'язана з гістонами одинарна двониткова суперспіралі- зована ДНК. Дві дочірні молекули ДНК, що знаходяться у двох хроматидах, утримуються разом у ділянці центромери за допомогою нереплікованого сегмента ДНК. Будова хромосоми на різних ділянках неоднакова. У хромосомах розрізняють первинну перетяжку, що поділяє хромосому на два плеча. Первинна перетяжка (центромера) - найбільш спіралізована частина хромосоми. Центромера являє собою загальну, нерепліковану ділянку ДНК. На ній розташовуються спеціальні білки, що утворюють кінетохори (від грец.χϊνησις, - рух, χώρα - місце), до яких при розподілі генетичного матеріалу прикріплюються нитки веретена. Це сприяє поділу дочірніх хроматид під час анафази. Місце розташування первинної перетяжки в кожній парі хромосом індивідуальне і стале, що зумовлює, головним чином, її форму. Кінці плечей хромосом одержали назву теломерів. Вони містять тисячі повторюваних послідовностей, наприклад, ТТАТТГ. Це генетично неактивні спіралізовані ділянки, що перешкоджають з'єднанню хромосом між собою або з їх фрагментами. Позбавлена теломери хромосома виявляється "липкою" і легко з'єднується з такими ж ділянками інших хромосом. Отже, теломери зберігають хромосому як індивідуальну дискретну одиницю, забезпечують її індивідуальність. Деякі хромосоми мають вторинні перетяжки, що часто відокремлюють ділянки хромосом, названі супутниками. Ці ділянки хромосом містять гени рРНК. Такі хромосоми в ядрах клітин людини можуть наближатися одна до одної, вступають в асоціації, що сприяє формуванню ядерець. Ці ділянки у хромосомах називають ядерцевими організаторами. У людини вторинні перетяжки є на кінцевих ділянках коротких плечей 13-15 і 21-22 пар хромосом, а також на довгому плечі 1-ї пари хромосом. Хромосома на стадії ранньої профази набагато довша і тонша, ніж метафазні хромосоми, тому в них можна розпізнати більше сегментів (рис. 1.41). Показані на рисунку сегменти виявляються шляхом фарбування препаратів барвниками, що зв'язуються переважно з А-Т - насиченими послідовностями ДНК. На рисунку представлено тільки по одній хромосомі з кожної гомологічної пари. Аутосоми та гетерохромосоми. Було відмічено, що практично в будь-якому каріотипі існує одна пара хромосом, що істотно відрізняється від інших. У самок ця пара була однакова, але різна в представників протилежної статі (самців). Хромосома, що була присутня двічі в парі в каріотипі самки й один раз у самця, була названа як Х-хромосома. Друга хромосома в самця була названа У-хромосомою. X- і Y -хромосоми називаються статевими хромосомами, або гетерохромосомами. Інші хромосоми з даного набору, що є однаковими для обох статей, називаються аутосомами. Чоловіки мають X- і Y -хромосоми і 22 пари аутосом. Жінки мають дві Х-хромосоми і 22 пари аутосом. Статеві хромосоми розподіляються при мейозі так само, як і інші хромосоми. Таким чином, кожний сперматозоїд одержує тільки одну статеву хромосому: або X, або Y. Оскільки чоловіча особина продукує два типи гамет, вона називається гетерогаметною. Жіноча особина продукує тільки один тип гамет, що мають тільки Х-хромосому, і називається гомогаметною. Стать майбутньої дитини визначається під час запліднення яйцеклітини. Якщо яйце запліднене сперматозоїдом, що містить Y -хромосому, зигота буде мати X- і Y -хромосоми і дасть початок розвитку чоловічої особини. Якщо яйце запліднене сперматозоїдом, що містить Х-хромосому, то зигота буде мати дві Х-хромосоми, що зумовить розвиток жіночої особини. Значення і функції хромосом. Хромосоми утворюються з хроматину на початку мітозу. Впродовж поділу клітини вони зазнають ряду структурних змін і знову утворюють хроматин в ядрах дочірніх клітин. Біологічне значення утворення хромосом та їх перетворень - рівномірний розподіл спадкового матеріалу в клітини, що утворюються. Потрібно відзначити, що поділ хромосом - це лише спосіб розподілу ДНК на рівні частини і передачі його дочірнім клітинам. В інтерфазній клітині хромосоми виконують такі функції: 1) збереження спадкової інформації у вигляді суворої послідовності нуклеотидів ДНК; 2) контроль метаболізму шляхом регуляції утворення необхідних ферментів; 3) забезпечення росту клітин, підтримка їхньої структури і функцій шляхом керування синтезом структурних білків; 4) контроль клітинного диференціювання під час розвитку; 5) забезпечення умов подвоєння ДНК. У клітинах, що діляться, утворюються візуально помітні хромосоми, що необхідні для таких функцій: 1) компактизація (ущільнення) спадкового матеріалу в тисячі разів, при цьому генетичний матеріал "консервується" і втрачає спроможність до експресії; 2) утворення структур, зручних для маніпуляцій при поділі (метафазна хромосома, що складається з двох хроматид); 3) рівномірний розподіл компактизованого неактивного генетичного матеріалу між дочірніми клітинами у вигляді окремих хроматид; 4) декомпактизація (розпушення) спадкового матеріалу й утворення активного інтерфазного хроматину.