Косміди
Косміди (Cosmides) — плазміди, що містять фрагмент ДНК фага лямбда включно зі cos-ділянкою. Разом із системами пакування у фагові частинки in vitro використовуються як векторні молекули для клонування генів та при побудові геномних бібліотек. Косміди вперше сконструювали Коллінс та Брюнінг 1978 року. Їхня назва походить від скорочення двох термінів: cos-ділянка (сам термін у свою чергу походить від англ. cohesive ends — липкі кінці) та плазміда.
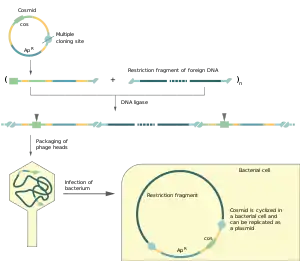
Косміда має розмір приблизно 5 т.п.н. і обов'язково включає такі елементи: cos-ділянку, необхідну для запаковування у віріони фага лямбда, точку ori плазміди для реплікації у клітині бактерії, сайти розпізнавання ендонуклеазами рестрикції та маркерний ген (наприклад ген резистентності до якогось антибіотику).
За допомогою космід можна клонувати ділянки ДНК розміром 32-47 т.п.н.. Для цього косміду та чужорідну ДНК обробляють однією і тією ж ендонуклеазою рестрикції, після чого отримані лінійні фрагменти змішують і проводять реакцію лігування. Далі відбувається пакування ДНК у фагові голівки in vitro, вона розрізається у cos-ділянках. Цей процес супроводжується селекцією фрагментів за розміром, оскільки запаковуватись може тільки ДНК довжиною 78—105% від геному фага лямбда. Таким чином у віріони потраплять переважно рекомбінантні косміди, що містять клоновану ділянку ДНК. Продукти лігування між двома космідами будуть замалими для пакування, а між двома фрагментами чужорідної ДНК — завеликими.
Отримані фагові частинки із рекомбінантними космідами використовують для інфікування клітин кишкової палички. У цитоплазмі косміди циклізуються завдяки з'єднанню липких кінців і далі реплікуються як звичайні плазміди, не проявляючи ніяких функцій фага лямбда. Селекція трансформованих клітин проводиться за маркерним геном присутнім у векторній молекулі.
Модифікації
Оскільки косміди дають можливість клонувати відносно великі ділянки ДНК, вони є зручними векторами для створення бібліотек фрагментів геномів еукаріот отриманих шляхом часткового розщеплення ендонуклеазами рестрикції. Проте продукти такого розщеплення можуть мати різний розмір, і можливі випадки, коли відбувається лігування двох відносно невеликих фрагментів, що не знаходились поруч у геномі, і створення відповідного клона, який даватиме хибне уявлення про їх положення в хромосомах. Цю проблему можна подолати шляхом фракціонування продуктів часткового розщеплення за розміром. Проте навіть такий метод не дозволяє повністю уникнути виникненню клонів космід, що містять кілька лігованих фрагментів ДНК, тому були запропоновані інші методи, зокрема дефосфорилювання фрагментів чужорідної ДНК, щоб вони не могли об'єднуватись одне з одним. Також були створені спеціальні космідні вектори pJB8 (Burke, Ish-Horowicz, 1981) та c2XB (Bates, Swift, 1983), при використанні яких імовірність утворення хибних клонів зводиться до мінімуму.
Сучасні косміди серій pWE та sCos характеризуються наявністю кількох сайтів клонування, фланкованими фаговими промоторами та унікальними ділянками розпізнавання ендонуклеазами рестрикції NotI, SacII та SfiI, що дозволяють вирізати вставку одним фрагментом.
Джерела
- Primrose S.B., Twyman R.M., Old R.W. (2002). Principles of Gene Manipulation (вид. 6th). Wiley-Blackwell. ISBN 0632059540.
- Griffiths A.J.F., Wessler S.R., et al (2004). An Introduction to Genetic Analysis (вид. 8th). W. H. Freeman. ISBN 978-0716749394. Архів оригіналу за 26 лютого 2015. Процитовано 20 квітня 2012.