Стратегії рослин
Стратегії рослин (англ. plants strategies) — способи виживання і підтримки стабільності популяцій в угрупованнях і екосистемах. Існує кілька систем типів стратегій. Так, Ерік Піанка (англ. Eric Pianka) розрізняє два типи, які називає r- і K-стратеги[1]. У першому випадку виживання досягається за рахунок витрати більшої частини фіксованої енергії на розмноження. у другому — на підтримку вегетативних процесів (наприклад, однорічники-рудерали і дерева).
Типи стратегій
Роберт Віттекер (англ. Robert Whittaker) виділяє три типи стратегій по динаміці чисельності популяцій між верхньою і нижньою межами їхнього існування[2]: К-стратеги підтримують чисельність біля верхньої межі за рахунок диференціації екологічних ніш, L-стратеги підтримують чисельність біля нижньої межі за рахунок переживання стресів у стані спокою (включаючи і пустельні однорічники, які можуть тривалий час зберігатися в стадії насіння) і r-стратеги — види чисельність популяцій яких коливається від верхньої до нижньої меж.
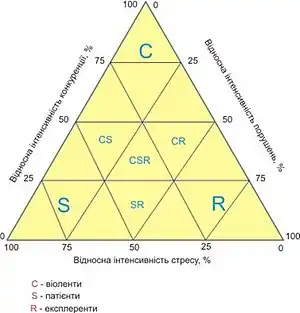
Л. Г. Раменський розрізняв віоленти, патієнти і експлеренти[3], що було практично повністю повторено пізніше в популярній системі стратегічних типів Дж. Грайма (J. Philip Grime), який представив її у формі трикутника, в кутах якого поміщені K-страгеги (конкуренти-віоленти), L-стратеги (стрес-толеранти-патієнти) і r-стратеги (експлеренти)[4].
Оскільки Дж. Грайм не розрізняв біотичного та абіотичного стресу, а групу рудералів трактував надмірно широко, відносячи до неї і пустельні малолітники, закономірно притаманні складу пустельних екосистем, і дають спалахи розвитку в сприятливі кліматичні періоди, за доцільне поєднавши системи Уїттекера і Раменського-Грайма розрізняти не три, а п'ять основних (первинних) стратегічних типів: віоленти (K) — види, здатні за рахунок активних біоморфологічних реакцій швидко захоплювати і утримувати за собою великий гіперпростір екологічних ніш; патієнти екотопічні (S) — види, здатні переносити абіотичний стрес і яким притаманний розвинений фізіолого-біохімічний апарат оптимального використання мізерних ресурсів (види пустель, тундри, галофіти, наскельні рослини і т. д.); патієнти фітоценотичні (SК) — види, які відчувають і пристосовані витримувати ценотичний стрес з боку конкурентів (тіньовитривалі види лісів, ацидофіли сфагнових боліт і т. д.); експлеренти несправжні (Rs) — однорічники пустель, що дають спалахи в період дощів, ефемери степів, ефемероїди лісів і т. д.; експлеренти справжні (R) — рудеральні рослини, пристосовані до життя в умовах збурень, види ранніх стадій сукцесії і т. д.
Виділені типи стратегій вельми рідко зустрічаються в чистому вигляді — природі більш властиві переходи між різними типами стратегій (вторинні типи стратегій), причому, відповідно до індивідуалістичної гіпотези стратегія кожного виду індивідуальна. Тому Дж. Грамм, виділивши в полі свого трикутника типи вторинних стратегій (RK, RS, SK, KSR), зображує різні види точками в полі цього трикутника, причому навіть представники однієї життєвої форми утворюють в цьому просторі деякі скупчення. Кожен вид, крім того, може змінювати тип стратегії в різних частинах свого ареалу, в різних угрупованнях, в різних умовах. Так. Ю. Р. Шеляг-Сосонко виділяє хоріони в межах ареалу дуба черешчатого (Quercus robur), де вид грає схожу фітоценотичну роль (тому, якщо в центрі ареалу вид повинен бути оцінений як К, то біля меж — як SK)[5]. Стратегії рослин близькі, але не тотожні фітоценотипам, адже останні показують тільки позицію виду в умовах досить сильної по внутрішньому ценотичному середовищу спільноти.
У цьому випадку група K еквівалентна едіфікаторам, група SK — ассектаторам. Однак, в умовах екстремумів, коли вплив популяцій рослин ослаблено і проявляються типи стратегій S і RS, фітоценотипи виділяти не можна і присвоєння більш рясним видам пустель рангу едифікаторів є грубою помилкою.
Крім стратегії підтримки особин в дорослому стані існують спеціальні типи стратегій розмноженні (перенесення насіння вітром, банк насіння, банк проростків і т. д.), які виявляються елементами загальної стратегії виживання.
Особливості r- і К-відбору і стратегій
| Характеристики | r-відбір і r-стратеги | К-відбір і К-стратеги |
|---|---|---|
| Клімат | Мінливий, непередбачуваний | Постійний, передбачуваний |
| Смертність | Катастрофічна, незалежна від щільності популяції | Викликана конкуренцією, залежна від щільності популяції |
| Крива смертності | Звичайно типу ІІІ | Звичайно типу І або ІІ |
| Розмір популяції | Мінливий, нерівноважний | Постійний, близький до граничної ємності середовища |
| Вільні ресурси | Поява вільних ресурсів, заповнення «екологічного вакууму» | Вільних ресурсів майже не буває, вони зайняті конкурентами |
| Внутрішньо- і міжвидова конкуренція | Слабка | Гостра |
| Розмір тіла | Відносно дрібний | Відносно крупний |
| Розвиток | Швидкий | Повільний |
| Статтева зрілість | Рання | Пізня |
| Швидкість розмноження | Висока | Низька |
| Розмноження протягом життя | Часто однократно | Неоднократно |
| Потомків у виводку | Багато | Мало, часто один |
| Кількість ресурсу на одного потомка | Низька | Висока |
| Тривалість життя | Коротка | Довга |
| Пристосування | Примітивні | Довершені |
| Оптимізується | Продуктивність | Ефективність |
Вторинні типи стратегій
Багатьом видам притаманні вторинні стратегії, тобто вони поєднують ознаки синдромів двох або трьох первинних типів стратегій: SR, CR, CS, CRS. Однак оскільки синдроми віолентності, патієнтності і експлерентності пов'язані трейдоффом[6], а величина «сумарного адаптивного потенціалу» обмежена, жоден вид з вторинною стратегією не може мати повний набір ознак двох і тим більше трьох первинних стратегій.
Це нагадує ситуацію із фондовим портфелем: в його складі можуть бути акції однієї або кількох компаній, але їх загальна вартість визначається величиною капіталу. У видів з вторинними типами стратегій співвідношення обсягів реалізованої та фундаментальної ніш може змінюватися від нуля до одиниці залежно від того, в яких співвідношеннях знаходяться властивості віолентності, патієнтності і експлерентноісті (чим вища частка експлерентності, тим воно ближче до нуля).
Видів рослин з вторинними типами стратегій більше, ніж видів з первинними типами стратегій. Приклад виду, що має стратегію віолент-патієнт (CS), є сосна, яка добре росте на бідних піщаних ґрунтах, а також всі види роду ялина, які зростають в холодному кліматі на бідних кислих (але добре зволожених) ґрунтах. І сосна, і особливо ялина є сильними домінантами, міцно утримують свої реалізовані ніші.
Стратегію віолент-рудерал (CR) мають такі види, як вільха сіра (Alnus incana), яка розростається на вирубках, і кропива дводомна — звичайний домінант ґрунтів, багатих азотом. Види зі стратегією патієнт-рудерал-патієнт (SR) досить рідкісні. Їх приклад — види роду Ребрик (Peganum), які можна спостерігати на витоптаних майданчиках навколо колодязів в пустельній зоні. Нечисленність видів зі стратегією SR є причиною нестійкості екосистем екстремальних умов до порушень, які викликаються господарською діяльністю людини. Відновлення рослинності аридних або арктичних пустель після порушень відбувається дуже повільно і, як правило, не повністю.
Більшість лугових і степових рослин представляють змішаний тип стратегії — CRS, тобто поєднують у своїй поведінці риси віолентності, патієнтності і експлерентності, хоча ці якості у різних видів представлені в різному співвідношенні. Наприклад, у видів солончакових луків — ячменю короткоостого (Hordeum brevisubulatum), покісниці розставленої (Puccinellia distanas) або типових домінантів степів — ковили і типчака — більше ознак патієнтності, а у пирія повзучого (Elytrigia repens), стоколосу безостого (Bromus inermis) і війника наземного (Calamagrostis epigeios) — експлерентності.
Пластичність стратегій
Багато видів рослин мають здатність у різних умовах використовувати різні ознаки свого адаптивного комплексу. У тварин ця властивість не виражена. Класичним прикладом, який ілюструє пластичність стратегії, є болотна форма сосни звичайної, яка представляє низку форм від високих дерев до карликових чагарникових форм висотою 60-80 см. Довжина хвої у «ліліпутів» не перевищує 1 см, а діаметр «стовбура» — становить 5-7 мм. Тривалість життя цих «дерев» досягає 100 років, на них утворюються шишки зі схожим насінням (іноді на одному «дереві» — всього одна шишка). З цього насіння в сприятливих умовах виростає звичайна високостовбурна сосна. Пригнічення сосни пов'язано з впливом наростаючого вгору торф'яного субстрату, який хоронить її кореневу систему. Коріння опиняються в умовах не тільки вкрай низької забезпеченості елементами мінерального живлення, а й кисневого голодування і холоду. Сфагнум є гарним теплоізолятором, і тому крига на глибині 20 см може зберігатися практично протягом усього літа.
Дуб звичайний в місцепроживаннях з оптимальними умовами — типове дерево-віолент, а на південній межі ареалу він представлений чагарниковою формою і є патієнтом.
Патієнтом на засолених ґрунтах є очерет, який в цих умовах представлений сланкою формою з вузькими листям. У плавнях дельт південних річок (Волга, Дон, Дніпро, Урал, Сирдар'я, Амудар'я та ін.) в умовах достатку елементів мінерального живлення і теплого клімату цей же вид має стратегію справжнього віолента, його висота досягає 3 і навіть 4 м, а ширина листка — 10 см. При цьому, очерет, як і сосна, утворює повний ряд переходу від віолентної високої форми до патієнтної карликової. Це можна спостерігати в дельті Волги в озерах-ільменях з різною солоністю води. Відмінності солоності води пов'язані з тим, що озера відчувають різний вплив річки. В озерах, які регулярно промиваються повеневими водами, вода прісна, а в тих, що промиваються періодично — солона. Озера, що зовсім втратили зв'язок з річкою, висихають і перетворюються на злісні солончаки.
Див. також
Література
- Пианка Э. Эволюционная экология. — М.: Мир, 1980. — 399 с.
- Работнов Т. А. Фитоценология. — М.: Изд-во МГУ, 1978. — 384 с.
- Раменский Л. Г. Введение в комплексное почвенно-геоботаническое обследование земель. — М.: Сельхозгиз, 1938. — 620 с.
- Раменский Л. Г. Избранные работы. Проблемы и методы изучения растительного покрова. — Л.: Наука, 1971. — 334 с.
- Уиттекер Р. Сообщества и экосистемы. / Пер. с англ. — М.: Прогресс, 1980. — 328 с.
- Шеляг-Сосонко Ю. Р. Ліси формації дуба звичайного на території України та їх еволюція. — К.: Наук. думка, 1974. — 240 с.
Праці Дж. Грайма
- Grime J.P. & Pierce S. The Evolutionary Strategies that Shape Ecosystems. Wiley-Blackwell. (2012) ISBN 0-470-67481-4
- Vegetation classification by reference to strategies. Nature (1974) 250: 26-31.
- Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist (1977) 111: 1169—1194.
- Plant Strategies and Vegetation Processes. Wiley. (1979) ISBN 0-471-99692-0
- Plant Strategies, Vegetation Processes, and Ecosystem Properties. (2nd much expanded edition of the above) Wiley (2001) ISBN 0-471-49601-4
- Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology (1998) 86: 902—910.
- Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences. Journal of Vegetation Science (2006) 17: 255—260.
- Plant strategy theories: a comment on Craine (2005). Journal of Ecology, 95, 227—230.
Виноски
- Пианка Э. Эволюционная экология. — М.: Мир, 1980. — 399 с.
- Уиттекер Р. Сообщества и экосистемы. / Пер. з англ. — М.: Прогресс, 1980. — 328 с.
- Раменский Л. Г. Избранные работы. Проблемы и методы изучения растительного покрова. — Л.: Наука, 1971. — 334 с.
- Grime J.P. & Pierce S. The Evolutionary Strategies that Shape Ecosystems. Wiley-Blackwell. (2012) ISBN 0-470-67481-4
- Шеляг-Сосонко Ю. Р. Ліси формації дуба звичайного на території України та їх еволюція. — К.: Наук. думка, 1974. — 240 с.
- Trade-off — зворотній зв'язок