Часова стала (нейрофізіологія)
Часова (або мембранна) стала - константа, яка описує вплив електричних властивостей мембрани нейрона на проходження по ньому електричного сигналу. Позначається грецькою літерою (тау).


Використання
Опис кінетики електричного потенціалу клітинної мембрани
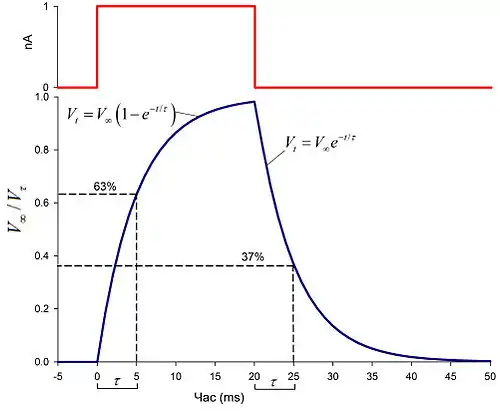
Наслідком електричних властивостей клітинної мембрани є те, що при проходженні через неї електричного струму її електричний потенціал змінюється не миттєво. Наприклад, при поданні ззовні прямокутного імпульсу струму (див. Рис. 1) мембрана поступово деполяризується протягом кількох мілісекунд, а потім так само поступово реполяризується після закінчення імпульсу. Таке уповільнення реакції виникає завдяки тому, що клітинна мембрана в розчині електролітів (цитоплазма та зовнішньоклітинне середовище) набуває властивостей електричного конденсатора. Для ідеалізованого випадку клітини, мембранний потенціал якої є однаковим в усіх точках поверхні, значення мембранного потенціалу V в деякий момент часу t (Vt) обчислюється згідно з рівнянням

протягом проміжку часу, коли мембранний потенціал зростає, і

протягом проміжку часу, коли потенціал знижується, тобто після закінчення імпульсу струму.
В цих рівняннях t - час, що пройшов після початку імпульсу (розмірність - мілісекунди), - часова або мембранна стала (розмірність - мілісекунди), V∞ (розмірність - мілівольти) - стале максимальне значення мембранного потенціалу, що може бути обчислене як
- ,

де rm - електричний спротив мембрани в мегаомах, І - сила струму в пікоамперах.
Часова стала також залежить від електричних властивостей нервової клітини, і ця залежність може бути описана як
- ,

де cm - електрична ємність клітинної мембрани в пікофарадах. При цьому значення rm та cm до великої міри залежать від розміру клітини: клітини великого розміру звичайно мають низькі значення rm та високі значення cm, і навпаки; з огляду на це в нейрофізіології електрична ємність мембрани часто використовується для порівняння відносних розмірів клітин.
Фізичний смисл часової константи , таким чином, це проміжок часу, при зростанні мембранного потенціалу (Vt) достатній для досягнення ним величини 1-1/е від V∞, або 63%; а при його падінні - величини 1/е від V∞, або 37% (див. Рис. 1). Тобто, чим більшим є значення мембранної сталої, тим повільніше проходить зміна електричного потенціалу клітини.
Опис кінетики електричного струму крізь клітинну мембрану
Аналогічно попередньому випадку, мембранна стала використовується для характеристики електричного струму, що генерується нервовою клітиною у відповідь на збудження. При цьому фізичний смисл - час, необхідний для досягнення 63% від максимального значення згенерованого струму (при його зростанні), або 37% максимального значення - при його падінні (див. Рис. 2). В останньому випадку ще називають константою деактивації рецепторів (якщо зникнення струму пов'язане із дисоціацією нейромедіатора від рецептора), або константою десенситизації (якщо нейромедіатор лишається зв'язаним з рецепторами, але вони, тим не менше, припиняють генерацію струму завдяки десенситизації).
Джерела
- Hodgkin, A. L. snd W. A. H. Rushton (1938) The electrical constants of a crustacean nerve fibre. Proc. R. Soc. Lond. 133: 444–479.
- Johnston, D. and S. M.-S. Wu (1995) Foundations of Cellular Neurophysiology. Cambridge, MA: MIT Press.
- Rall, W. (1977) Core conductor theory and cable properties of neurons. In Handbook of Physiology, Section 1: The Nervous System, Vol. 1: Cellular Biology of Neurons. E. R. Kandel (ed.). Bethesda, MD: American Physiological Society, pp. 39–98.