Жаба гостроморда
Жаба гостроморда (Rana arvalis) — вид жаб роду Rana, поширений в Європі та Азії. Ця жаба характеризується однотонним забарвленням черева та великими плямами на вухах та часто (але не завжди) блідішою смужкою на спині. Досягає 7 см завдовжки, її голова більш витягнута, ніж у звичайної європейської жаби (Rana temporaria). Ноги частково перепончасті, задні ноги коротші, ніж у інших жаб. Інші назви: жаба мочарова, болотяна жаба.
? Жаба гостроморда | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Жаба гостроморда | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
| Rana arvalis (Nilsson, 1842) | ||||||||||||||||
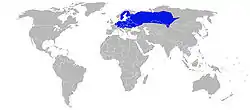 Ареал Жаби гостромордої Збережений (резидент) | ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||

Ареал розповсюдження
Ареал обіймає простір від північно-західної Франції, Бельгії, Нідерландів, Німеччини,Швеції та прилеглих районів Норвегії,Фінляндії на південь до Альп, Хорватії, Словенії, частково Італії, північної Румунії та на схід до Якутії, Алтаю (Росія), доходить до північного Китаю.
В останньому зведенні про поширення європейських земноводних та плазунів з двох підвидів наводиться лише один — R. а.wolterstorffi — і вказано, що він трапляється у південно-східній Польщі, Австрії, Угорщині, Румунії, Словенії, Хорватії та у Західній Україні (таксономічний статус гостромордих жаб в інших частинах Європи не вказаний).
Згідно з більшістю сучасних європейських досліджень цього виду, в Україні живуть номінативний підвид та R. а. wolterstorffi (довгонога або угорська жаба). Виособлення R. arvalis від спільного стовбура, сформованого разом з іншим спорідненим видом (R. temporaria), відбулося на межі раннього й пізнього пліоцену близько 3 млн років тому, а наступне формування двох підвидів (R. a. arvalis та R. a. wolterstorffi) здійснилося вже близько 1 млн років тому в плейстоцені.
В Україні амфібії цього виду виявлені у ранньоголоценових покладах з Тернопільської області, хоча є докази про наявність цих тварин у пліоцен-голоцені на території теперішніх Черкаської та Херсонської областей.
В Україні гостроморда жаба живе на її більшій частині і прив'язана до лісової та почасти лісостепової зон; у степовий регіон заходить лише долинами великих річок — до пониззя Дніпра (околиці Голої Пристані), Південного Бугу (середня течія р. Чичиклія), є вона й у дельті Дунаю. У Криму відсутня. У Карпатах трапляється лише на рівнинах та в передгір'ї, у гори ж може проникати річковими долинами, але звичайно не вище 1000 м.
Типові стації — більш відкриті та сухі біотопи, ніж у трав'яної жаби. Це ліси, сухі бори, рідколісся, галявини, узлісся, луки, береги боліт та річок тощо. В горах відомі знахідки гостромордих жаб на висоті 800 м над рівнем моря.
Гостроморда жаба майже скрізь пов'язана з деревно-чагарниковими насадженнями, прилеглими до боліт, лук, галявин, лісових вирубок, полів тощо. Порівняно з трав'яною жабою R. arvalis характеризується більшою синантропністю, нерідко трапляється на дачних ділянках, у селищах, у приміській зоні, у парках, садах. Разом з тим цей вид при виборі місць розмноження віддає перевагу природним, а не антропогенним водоймам.
Чисельність
При порівнянні різних типів водойм, що їх обирали гостроморді жаби, встановлено, що в калюжах, утворених на місцях роботи бульдозера, кладок R. arvalis не було; у схожих водоймах, але більш раннього походження і з зарослим дном (старі кар'єри) виявлено 4 кладки, а у природному — 18 кладок (околиці м. Сиктивкар, АР Комі, Росія).
Дані з чисельності гостромордих жаб у різних регіонах України фрагментарні. Судячи з наявних матеріалів із Закарпаття, гостроморді жаби віддають тут перевагу світлим і уникають хвойних лісів, чисельність цього виду становить від 7-11 ос./1000 м до 5-20 ос./100 м маршруту.
Слід підкреслити, що чисельність R. arvalis суттєво залежить як від типу біотопу, так і від сезону активності. Наприклад, навесні у долині Південного Бугу щільність гостромордих жаб становить 85-97 ос./га у водоймах, прилеглих до полів, 67-76 ос./га у прилеглих до лісу, 53-60 ос./га в чагарникових ділянках, 21-24 ос./га на лісових вирубках і 11-14 ос./га в лісах. Восени у цьому ж регіоні чисельність цього виду в чагарниках досягає 18-22 ос./га, на лісових вирубках до 40-45 ос./га, а у лісах 26-30 ос./га.
На дніпровських островах у районі Херсона на початку жовтня чисельність R. arvalis на 100 м маршруту становить 15-20 особин. У долині річки Орель (Дніпропетровська обл.) їхня чисельність в сезон активності коливається у різних типах біотопів від 0,3 до 6,1 ос./10 циліндро-діб, по закінченні сезону розмноження кількість гостромордих жаб як у заплавних лісових ділянках, так і на відкритій призаплавній луці досягає 5-10 ос./100 м маршруту. У звичайних біотопах цього ж регіону (степове Придніпров'я) чисельність гостромордих жаб становить 19,0-53,0 ос./га, хоча у деструктивних місцях перебування (рудно-вугільні відвальні техногенні ландшафти) щільність знижена до критичного рівня 0,6-3,1 ос./га.
Щільність амфібій цього виду за межами української частини ареалу (Карелія, Архангельська обл.) коливається від 4 ос./га, у заплаві р. Північна Двіна до 0,19-0,20 ос./га у сосняках чи темнохвойних лісах. R. arvalis у вторинних сосново-березових лісах Печоро-Іличського заповідника (північна тайга європейського північного сходу Росії) трапляється на 1 км маршруту чисельністю від 0,4 до 1 ос. Дані обліків залежать від сезону активності: при обліку 17 травня на маршруті 8 км зареєстровано 92 особини, а 1 червня — лише одна. На іншій ділянці цього регіону щільність у червні була 6 ос./100 м², у серпні, після виходу цьоголітків — 15 ос./100 м² (Ануфриев и др., 1996).
У східніших популяціях (Приєнисейська тайга), чисельність гостромордих жаб у лісах першої надзаплавної тераси становить 9, а на другій 2-3 ос./100 циліндродіб, хоча у більш південних тайгових ділянках вона може бути в 3-3,5 разу більше. На південному сході Західного Сибіру її чисельність у низинних болотах досягає 597,5 ос./га, а на луках центральної заплави р. Об — 454,4 ос./га.
Зовнішній вигляд
Тварини середніх розмірів, довжина тіла статевозрілих особин 40-64 мм. Дослідження морфологічних ознак тварин із Закарпаття показало, що довжина тіла самців становить близько 52 мм, самиць — 53 мм. Внутрішній п'ятковий горб високий. Голова загострена, на спині є дві поздовжні спинно-бокові складки, у забарвленні переважають коричнювато-світлі або оливково-бурі тони, позад голови добре виявлена темна L-подібна пляма, черевний бік без плям. По боках голови розташовані темні смуги, які доходять до основи передніх кінцівок.
Існують експериментальні дані (які потребують, вірогідно, перевірки й підтвердження), що ознаки крапчастості та плямистості є рецесивними по відношенню до напівкрапчастості й напівплямистості. Задні кінцівки розвинені добре і, якщо їх витягнути уздовж тіла (у напрямку до голови), то скоковий (надступаковогомілковий) суглоб може доходити до очей (представники номінативного підвиду), а в інших — дещо заходить за кінець морди (жаби довгоногого підвиду wolterstorffi).
На задніх лапах часто є візерунок з декількох поперечних плям-смуг, пальці тут з'єднані плавальною перетинкою. Позад очей розташована добре виявлена барабанна перетинка. Самці мають внутрішні резонатори, в них розвинені шлюбні мозолі на перших пальцях передніх кінцівок.
Статевий диморфізм у забарвленні особливо добре помітний в сезон нересту і виявляється у тому, що для самців характерна поява загального блакитного тла. Відомості про статевий диморфізм у розмірних показниках суперечливі. Так, вони відсутні в гостромордих жаб із Закарпаття, але виявлені при аналізі морфологічної мінливості представників цього виду в інших регіонах України.
Мінливість

У зведенні з батрахофауни Карпат R. a. wolterstorffi вказана для Закарпаття. Однак наступне порівняння двох діагностичних показників, запропонованих автором опису цього підвиду (співвідношення L./T. і T./L. t. ci.), показало поширення «довгоногих» жаб не лише у Закарпатті, але й у ряді інших регіонів України, як, втім, і популяцій, що характеризуються проміжними (у порівнянні з «коротконогими») значеннями. Така специфіка просторової мінливості цих ознак (брак чітких географічних меж у поширенні кожного морфологічного типу), не дозволила дійти остаточного таксономічного висновку. Не виключено, що та ж сама причина стала основою для твердження про перебування в одному й тому ж самому місці у Закарпатті (Долина нарцисів) різних «морфологічних типів»: «arvalis», «dalmatina» та «гібридів». Після ідентифікації генетичними методами була показана приналежність усіх їх до R. dalmatina.
Разом з тим ситуація, коли тварин з ознаками номінативного підвиду виявляли в районах поширення підвиду wolterstorffi відзначалася раніше й на території Австрії. У північно-східній Україні, за останніми даними, трапляються R. a. wolterstorffi та R. a. arvalis, однак вони прив'язані до різних місць перебування. Перші поширені уздовж степових ділянок та в деструктивних біотопах (включно з сосновими лісами) поблизу великих річок, тоді як другі проникають з Росії нативними (одвічними, первинними) листяними лісами. У досліджуваному регіоні обидві форми симпатричні. Поки що невідомо, чи є між ними генетичні відмінності, чи це екологічні раси, своєрідність яких базується на епігенетичному феномені.
Географічна (міжпопуляційна) мінливість морфологічних ознак у межах України вивчена слабо. На Закарпатті відзначені деякі відмінності в розмірних показниках самців та самиць (наприклад індекс L./T. в самців 1,80-2,12 і 1,82-2,14 в самиць), однак вони не достовірні. Разом з тим популяціям цього виду з інших частин України властиві достовірні відмінності за цим показником і самці характеризуються відносно довшими задніми кінцівками. Так, у самців цей індекс коливається в межах 1,72-1,88, а в самиць 1,82-2,0. Самцям також властиві дещо більші розміри тіла (45,18-53,99 мм самці та 45,18-52,62 мм самиці).У гостромордих жаб з території України та з Литви цей індекс у представників обох статей коливається в межах 1,71-2,21.
Спроби з'ясувати залежність деяких розмірів від факторів зовнішнього середовища дозволили стверджувати, що темп (індекс) росту гомілки більший в умовах теплого клімату.
Для представників цього виду справді характерний рідкісний випадок статевого диморфізму, при якому самці мають більшу довжину тіла порівняно з самицями. Такий характер статевого диморфізму відрізняється від іншого близького виду (трав'яної жаби) тим, що ці відмінності спостерігаються у всіх вікових групах R. arvalis, а в R. temporaria розміри самиць починають перевищувати розміри самців лише після 3-річного віку.
Порівняння морфологічної диференціації R. arvalis з сусідньої Білорусі показало, що жаби з довшими ногами живуть у її підвищеній частині, а тварини з південних регіонів відрізняються ширшею мордою, зменшеними очима та барабанною перетинкою.
З іншого боку, географічна мінливість індексу F./T. на ділянці між 50°-60° пн.ш. і 57,5° та 67,5° сх. д. найскоріше відсутня. Разом з тим серед статевозрілих самців існує зростання в північному напрямку індексу довгоногості при великій кількості локальних відхилень. Порівняння ознак черепа жаб центральної та північної частин ареалу (Росія, Іванівська та Архангельська обл. відповідно) показало, що на півночі R. arvalis більші.
Слід зазначити, що специфіка личинкового розвитку може справляти помітний вплив на фенотип окремих генерацій. Так, підвищена щільність утримання пуголовків приводить не лише до дрібніших розмірів цьоголітків, але ай до суттєвих відмін (у порівнянні з контролем) за ознаками L./T. і F./T.
R. arvalis з Угорщини відрізняється від представників цього виду у Польщі та Румунії довшими задніми кінцівками, хоча статистичний аналіз показав низький відсоток правильної класифікації вибірок. Крім того, не зважаючи на те, що всі вибірки з Польщі та Румунії віднесені до номінативного підвиду, а вибірки з Угорщини — до R. a. wolterstorffi, з'ясувалося, що використання індексів L./T. і T./L. t. ci. усе ж ненадійне для ідентифікації цих підвидів.
В околицях Києва частота розподілу морфи «unicolor» (особини, позбавлені візерунка) порівняно низька й приблизно така сама, як і в степовій частині південного Уралу і на півдні півострова Ямал (2,3-8,3%). Частота морфи «striata» (серединна світла смуга) в околицях Києва становить 5,1-6,5%, що відповідає даним з долини р. Ангара у Східному Сибіру. У Білорусі зона поширення жаб морфи «striata» обмежена прикордонним з Україною басейном річки Прип'ять.
Разом з тим, у північному напрямку частота трапляння морфи «striata» знижується: 85% (51° пн.ш.) — 36% (57° пн.ш.) — 22% (67° пн.ш.). Судячи з останніх досліджень, більша частота цієї морфи пов'язана з відкритими біотопами. Серед проаналізованих у межах всього ареалу гостромордих жаб для 42,4% характерним є фенотип М, hM — 19,4%, B — 13,2%, P — 1,25%, S — 37%, достовірні відміни в особливостях забарвлення між самцями й самицями виявлені лише в 3 з 21 досліджених популяцій.
Не виключено, що якась частка в загальній мінливості зовнішньої морфології пов'язана з декількома факторами. На урбанізованих ділянках у Росії відсоток особин з депігментованою райдужкою в цьоголітків доволі високий (5,7%), що, вірогідно, було наслідком близькоспоріднених схрещувань та мутацій. При цьому є відміни між ювенільними самицями й самцями (серед самців істотно вищі). Частота відхилень у будові скелета серед самців також виразно більша, ніж серед самиць. Не виключено, що це явище пов'язане з загальною низькою мінливістю самиць та їхньою більшою онтогенетичною стабільністю. Крім того, у міському районі збільшена частота морфи «striata». Цікаво, що в цієї морфи поріг подразливості нервової тканини істотно нижчий, ніж у інших морф.
Генетика
Вивчення каріотипів бурих жаб України підтвердило 24-хромосомний диплоїдний набір R. arvalis (2n = 24, NF = 48), на коротких плечах другої пари хромосом іноді трапляються вторинні перетяжки.
Перетворення 26-хромосомного диплоїдного набору на 24-хромосомний відбувалося, найвірогідніше, двічі, при цьому перше зменшення кількості хромосом (шляхом злиття 11-ї пари з12-ю або з 13-ю) супроводжувало формування R. arvalis, а друге — решту 24-хромосомних таксонів групи бурих жаб (живуть за межами України).
Рівень гетерозиготності (Hexp) гостромордої жаби з Київської області становить 0,034, а генетичні дистанції між R. arvalis і R. temporaria досягають 0,260-0,294, між R. arvalis і R. dalmatina — 0,699-0,703,між R. dalmatina і R. temporaria — 0,613-0,651. Разом з тим генетична дистанція між гостромордою жабою з Люксембургу та трав'яною жабою з околиць С.-Петербурга істотно більша — 0,742. При цьому відмічено, що серед восьми 26-хромосомних видів (R. japonica, R. tsushimensis, R. okinavana, R. longictus, R. temporaria, R. asiatica, R. amurensis, R. latouchii) генетична дистанція коливається в межах 0,294-2,913, а серед чотирьох 24-хромосомних (R. ornativensis, R. chensinensis, R. dybowskii, R. arvalis) 0,260-1,018. Генетична ж дистанція між 26- і 24-хромосомними представниками була в межах 0,410-2,266.
Судячи з генетичних матеріалів та даних морфометричного аналізу (включно з остеологією R. temporaria, R. dalmatina, R. iberica, R. camerani, R. macrocnemis, R. holtzi, R. graeca, R. latastei), трав'яна жаба (разом з R. iberica) відділилася від предкового стовбура у пізньому пліоцені (близько 2,6 млн років тому). Прудка жаба (разом з R. graeca) сформувалася під час міндельського зледеніння четвертинного періоду (близько 700 тис. років тому).
Ще одні матеріали з генетичної диференціації південної та північної популяційних груп гостромордої жаби (перші живуть на низині Панонія, другі займають решту ареалу), свідчили про те, що дистанція Нея між гостромордими жабами з низини Панонія і цими амфібіями з решти ареалу становить 0,062, що дозволяє стверджувати, що останні зайняли свою ділянку ареалу відносно недавно.
Разом з тим, результати статистичного аналізу ознак зовнішньої морфології носять суперечливий характер, і крім того, ізольована популяція з Речі (Reci, Румунія), яка генетично належить до жаб з Панонії, займає проміжне становище.
Життя у природі
Весняна поява чимало залежить від особливостей погоди конкретного року, але зазвичай припадає на початок-середину березня. Найбільш ранні зустрічі на Закарпатті зареєстровані вже при температурі повітря 5 °C, хоча ці показники для басейну річки Південний Буг вище, і перші жаби відзначаються при температурі повітря 11,7 °C, води — 11,6 °C.
Парування починається відразу після весняного пробудження й появи на місцях розмноження. Для нересту гостроморда жаба звичайно обирає лісові ставки й калюжі. Свіжовідкладена ікра спочатку плаває у товщі води на глибині 15-20 см, а потім спливає на поверхню. Кладка ікри в самиць з довжиною тіла близько 52 мм становить приблизно 730 штук, при довжині тіла близько 73 мм — понад 2600. Для тварин цього виду характерне групове відкладання ікри: звичайно в одній водоймі відкладають ікру десятки пар.
Загальна тривалість нересту в одній водоймі становить від 3 до 28 діб. Цей показник на урбанізованих та неурбанізованих територіях у різних ділянках ареалу відрізняється. На південному сході Західного Сибіру в кладках на урбанізованих територіях було близько 900 ікринок, на неурбанізованих близько — 1000, а в Підмосков'ї навпаки — відповідно 1682 і 1100.
У східній частині ареалу (Камське Передуралля) плодючість нижча й становить від 400 до 933 ікринок. У середній тайзі європейського Північного сходу Росії цей показник для гостромордих жаб при довжині тіла 45-53 мм становить 654 ± 39 шт., а при довжині 54-62 мм 906 ±80 шт. Період ембріонального розвитку на Закарпатті залежить від температурного режиму, та найчастіше становить два тижні, метаморфоз завершується приблизно за 3 місяці.
В умовах центрально-південних областей України тривалість личинкового розвитку становить близько 60 днів. У дещо північніших ділянках ареалу в Тамбовській області період водного розвитку становить приблизно 90 днів. Загальні розміри личинок після вилуплення 5-8 мм, перед проходженням метаморфозу — 15-48 мм. До появи передніх кінцівок личинки гостромордої жаби добре відрізняються від R. temporaria загостреним кінчиком хвоста, який у трав'яної жаби заокруглений.
Розміри цьоголітків відразу після метаморфозу за даними вивчення цього виду в басейні Південного Бугу становлять 7,7-15,7 мм. Особливо слід зазначити, що різні морфи (різні генотипи), наприклад «striata» і «maculata», хоча й ростуть в однаковому темпі, характеризуються відмінами у швидкості метаморфозу. Так, у різні роки в період з 1966 по 1971 р. чисельність «maculata» на околицях Челябінська становила від 55,3 до 72,2% серед усіх цьоголітків, що вийшли на сушу, досягаючи свого максимуму в посушливий рік.
Такі відмінності в чисельності можуть бути наслідком не лише неоднакової швидкості росту личинок з різним генотипом, але й більш раннім початком розмноження відповідних батьківських пар (генотип «maculata»). Мінливість частоти трапляння різних морф вірогідно пов'язана з дією антропогенного фактору, у всякому разі його посилення приводить до різкого зростання чисельності смугастих особин. Виживаність на антропогенних ділянках залежить від ступеня їх урбанізованості.
Так, на початкових та середніх етапах личинкового розвитку в зоні багатоповерхової забудови зберігається близько 0,92% усієї кількості відкладених ікринок (Єкатеринбург, Росія), в районі малоповерхової забудови — 11,9%, а в лісопарку — 24,2%. Підрахунок відкладених ікринок і кількість личинок у кінці водної стадії в забрудненій водоймі в околицях мідеплавильного заводу та у двох водоймах на відстані 6 і 15 км від джерела забруднення показав, що у першій водоймі збереглося 11,4% личинок, у другій — 1,6% і в третій — 10%.
Маса цьоголітків при метаморфозі становила близько 400, 329 і 562 мг відповідно (специфіка такої залежності потребує спеціального вивчення). Аналіз дії різних факторів показує, що на особливості біології цього виду (початок відкладання ікри, строки вилуплення пуголовків, метаморфоз тощо) істотний вплив справляє не тільки температура, але й, що має особливу вагу, попередні фенологічні явища, такі як температура лютого і березня, терміни танення снігу, червневі коливання температури і т. ін.
Гостроморді жаби пов'язані з водоймами лише в період нересту. По його закінченні за сховища на суші для них зазвичай правлять порожнини у старих пнях, ґрунті, листовій підстилці. Дані про добову активність обмежені. Так, на Закарпатті гостромордим жабам властива висока активність у денні та присмеркові години. У центральній Україні амфібії цього виду в сезон нересту активні цілодобово, а після його закінчення, за умов спекотної погоди — у вечірні години; в похмуру погоду або ж восени активні ранком та увечері. Це саме підтверджується відомостями про цих тварин у заплаві Дніпра (Київська, Дніпропетровська обл.).
У середній тайзі європейської частини Росії (басейн р. Вичегда) їхня найбільша активність відзначається з 20.00-22.00 до 2.00-4.00 годин. Гостромордим жабам властиві сезонні міграції. Так, мічення R. arvalis в околицях Львова дозволило встановити існування зворотних міграцій, при яких ці земноводні по закінченні розмноження полишають місця нересту та переміщуються на місця постійного перебування влітку.
Ці дані підтверджуються й схожими спостереженнями в інших ділянках ареалу. Так, у західній частині Німеччини дорослі тварини починають мігрувати на місця нересту ще восени й продовжують такі переміщення рано навесні. Після нересту вони покидали став і здійснювали зворотну міграцію на літні місця перебування. При цьому нестатевозрілим особинам (двохліткам) також властива весняна міграція на нерестові місця навесні і зворотна — на літні місця пробування.
Вивчення активності у місцях постійного літнього пробування на півночі європейської частини Росії показало доволі широку мінливість у рухомості: частину тварин реєстрували протягом 20 днів на ділянці всього 6 м², хоча інші за день переміщувалися до 150 м. Вибір місць для зимівлі мінливий. Так, у Передкарпатті (Львівська обл.) R. arvalis виявляли взимку на глибині 1,2 м у мулі незамерзаючої торф'яної ями, заповненої водою. У північній частині поширення цього виду (європейська частина Росії), зимівля тварин відбувається на суші, а у найпівнічнішому краї ареалу— у воді.
Багаторічні дослідження специфіки розмноження у різних частинах ареалу показали, що розміри тварин, що розмножуються вперше, становлять 38-55 мм. Строки досягнення статевої зрілості у різних ділянках ареалу відрізняються, але в середньому відповідають 2-5 рокам, а тривалість життя коливається від 6 до 11 років. Вже зазначалося, що тривалість життя R. arvalis в Україні істотно нижча, ніж у популяціях цього виду в інших частинах його ареалу, а відносна чисельність жаб віком понад 5 років невелика.
Частота трапляння особин старших вікових груп у різних регіонах відрізняється, і найстаріші особини частіше трапляються у високих широтах (тундра) або в горах (Алтай), де тварини 8чи 9-річного віку в популяціях звичайні. У цьому відношенні серед популяцій Білорусі, Південного Уралу, Московського регіону R. arvalis особини старші 5 років являють рідкість. До складу їжі личинок входять водорості, вищі рослини, детрит та істотно менше водних безхребетних.
Дорослі гостроморді жаби належать до числа видів, у складі живлення яких переважають наземні форми. За даними аналізу особливостей живлення цього виду на Закарпатті у раціоні переважають жуки (до 66%), плівкокрилі та двокрилі (до 16%), метелики (до 14%), простокрилі (до 10%) та дощові черви (до 18%). Загалом можна сказати, що спеціалізація у живленні відсутня, і їхній раціон, найвірогідніше, віддзеркалює відносну чисельність та доступність різних груп безхребетних у біотопах, населених амфібіями цього виду. Специфіка живлення R. arvalis з іншими типово наземними видами безхвостих амфібій почасти збігається, наприклад частка спільних ресурсів живлення з землянкою становить 88%.
Личинок гостромордих жаб виявляли у живленні старших за віком пуголовків трав'яних жаб. Після метаморфозу R. arvalis стає здобиччю багатьох видів риб (окунь, щука тощо), птахів (крижень, сіра чапля, річкова чайка, сіра ворона, грак, сорока і т. ін.), ссавців (кріт, видра, норки, сірий пацюк, кабан і т. ін.). Відзначені неодноразові напади на них медичних п'явок (Hirudo medicinalis), які можуть призводити до летального наслідку.
Охорона
Серед основних факторів, що справляють помітний вплив на скорочення чисельності та площі ареалу гостромордої жаби, слід передовсім вказати на руйнування природних біотопів цього виду під впливом антропогенного фактору. Вид занесено до Додатку II «Конвенції з охорони дикої флори і фауни та природного середовища існування в Європі» (категорія «види, що підлягають особливій охороні») та до «Червоної книги хребетних Міжнародного союзу охорони природи (МСОП)».
Примітки
- Kuzmin, S., Tarkhnishvili, D., Ishchenko, V., Tuniyev, B., Beebee, T., Anthony, B.P., Schmidt, B., Ogrodowczyk, A., Ogielska, M., Babik, W., Vogrin, M., Loman, J., Cogalniceanu, D., Kovács, T. & Kiss, I. (2009). Rana arvalis: інформація на сайті МСОП (версія 2009) (англ.) 14 грудня 2008
Посилання
Писанець Є. Земноводні України (посібник для визначення амфібій України та суміжних країн). — Київ : Вид-во Раєвського, 2007. — 197 с.
Ресурси Інтернету
- Rana arvalis МСОП
- Google Books. Concise Encyclopedia of Biology. Процитовано 27 березня 2006. (англ.)