Консорція (екологія)
Консорція (від англ. consortium — співучасть, співтовариство) — ценотична популяція вільноживучої зеленої рослини разом з усіма пов'язаними з нею популяціями інших організмів (тварини, бактерії, гриби і т.і.).

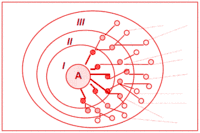
Гіпотеза консортивних зв'язків Беклємішева — Раменського — уявлення про існування основного осередка трансформації енергії в екосистемі, що є її структурною частиною і званою консорцією. Важливою відмінною рисою консорції є спільність «еволюційної долі», взаємне пристосування видів-консортів один до одного в процесі еволюції (коадаптаціі). Уявлення про консорції були незалежно сформульовані зоологом В. М. Беклємішевим в 1951 р.[2] і ботаніком Л. Г. Раменським в 1952 р.[3]
Великий теоретичний і методичний внесок у вивчення консорцій був зроблений естонським геоботаніком В. В. Мазингом (1966)[4]. Зокрема, він розрізняє індивідуальні, клональні, популяційні, регіональні, видові консорції. Ядром індивідуальної консорції зазвичай виступає автотрофна рослина-едіфікатор, компонентами (видами-консортами) є безпосередньо пов'язані з ним (трофічно і топічні) організми. Ядром популяційної консорції є вся популяція або вид в цілому, синузіальної консорції — види однієї екобіоморфи (наприклад, темнохвойні дерева). Крім того, В. В. Мазинг розрізняє консорції за числом трофічних рівнів-концентрів, на яких відбувається трансформація енергії (концентр автотрофів, фітофагів, зоофагів першого порядку, зоофагів другого порядку і т. д.). У міру підвищення рівня концентрації змінюється співвідношення його факультативних і облігатних елементів, що переводить консорцію з дискретних клітинок трансформації енергії в клас безперервних явищ.
Більшість дискусій щодо структури консорцій, активно велися в 1960-70-х роках, зводилося до вирішення трьох основних питань (Мальцев, 1987, с.47):
- Чи може бути детермінантом будь-який (автотрофний чи гетеротрофний) організм чи консорція пов'язана з автотрофною неепіфітною рослиною;
- Чи пов'язана консорція з окремою особиною детермінанта або з усією його популяцією;
- Якого характеру зв'язки слід відносити до консортивних (зокрема, чи слід дотримуватися уявлень про поліконцентричну структуру консорції, представлену на рис. 1, і чи відбиває це становище того чи іншого виду в харчовому ланцюгу).
На ці питання В. І. Мальцев (1987)[5] дає такі відповіді:
- Важливий не спосіб виробництва органічної речовини, а розміри організму і його здатність «контролювати» середовище (в цьому контексті роль автотрофів вище, хоча і не виключається консорція з детермінантом-гетеротрофом);
- Консорцію слід пов'язувати з ценопопуляцією детермінанта (популяційна консорція по В. В. Мазингу);
- Консортивні зв'язки розрізняються за своєю приуроченістю до ценопопуляції детермінанта [наприклад, манника водного (Glyceria maxima)] — власне консорт [личинки хірономід (масові види Glyptotendipes glaucus і Pentapedilum sordens)], супраконсорти (хижі п'явки роду Erpobdella, черевоногі молюски Lymnea stagnalis і Planorbarius corneus) і «відвідувачі» (представники нектону — риби).
Між детермінантом консорції і консортами існують різноманітні зв'язки, які, ґрунтуючись на результатах класифікації Т. А. Работнова (1994, 1996), можна звести до наступного (див. табл.
Таблиця
| Характер зв'язків | Інтерпретація |
|---|---|
| Вплив детермінанта консорції на консортів | |
| Фабричні | Дає консортам або опору (для ліан), або матеріал для побудови гнізд |
| Трофічні | Забезпечує консортів енергією чи речовинами, джерелом яких можуть бути живі органи детермінанта, його діаспора і пилок, прижиттєві виділення, відмерлі органи та ін. |
| Форичні | Запилення квітів та розповсюдження діаспор детермінанта консорції |
| Дефензивні | Забезпечує захист консортів від їхніх ворогів |
| Аттрактивні і репелентні | Виділяє речовини, які залучають або відлякують консортів |
| Поліпшення умов зростання | Бере участь в утворенні загальних умов проживання (мікроклімат, мікроґрунтові особливості тощо) |
| Вплив консортів на детермінанта консорції | |
| Фабричні | Використання детермінанта як місця і матеріалу для влаштування гнізд |
| Трофічні | Використання детермінанта як джерела енергії та елементів мінерального живлення. Поліпшення забезпечення детермінанта елементами мінірального живлення (фосфором — мікоризоутворюючі гриби, азотом — азотфіксуючі симбіонти); забезпепечення елементами мінерального живлення і, частково, енергією — комахоїдних рослин-детермінантів |
| Контактні | Травмування надземних і підземних органів (прогризання листя, обламування гілок, пошкодження кореневої системи тощо); іноді це супроводжується вегетативним розмноженням детермінантів |
| Асоціативні | Забезпечення детермінанта елементами мінерального харчування в результаті розкладання і мінералізації його відмерлих органів сапрофітними консортами |
| Форичні | Перенесення консортами пилку і діаспор |
| Патогенні | Виділення консортами-ендобіонтами токсичних метаболітів в тканини детермінанта |
| Едифікуючі | Зміна місця існування детермінанта в результаті діяльності консортів-тварин |
| Дефензивні | Захист детермінанта (яблуня — мурашки — попелиця) |
| Ареалографічні | Вплив консортів-запилювачів на межі розповсюдження детермінанта |
Консорти експлуатують окремі особини детермінанта, їх велика кількість в більшій мірі визначається фізіологічним станом детермінанта. Супраконсорти експлуатують уже ценопопуляцію детермінанта і, певною мірою, менше схильні до його детермінуючого впливу. Таким чином, консортивна сукцесія повинна йти від переважання топічних зв'язків над трофічними (превалювання супраконсортів над консортами) у напрямку до зростання трофічних відносин (зростання відносної частки власне консортів). Цей висновок підтверджується даними Л. М. Зимбалевської (1966), яка показала, що в міру становлення гідробіологічного режиму Кременчуцького та Київського водосховищ спостерігалося зменшення кількості прибережно-фітофільних видів (в переважній більшості — супраконсортів) і зростання відносної кількості фітофільних видів (в основному — консортів).
О. О. Корчагін (1976) і Т. О. Работнов (1983) розрізняють п'ять форм динаміки консорцій:
- Сезонну (пов'язану із сезонними змінами компонентів консорції).
- Флуктуаційну (різнорічні зміни чисельності та життєвого стану консортів).
- Сукцесійну (пов'язану з сукцесією рослинних угруповань).
- Онтогенетичну (пов'язану з онтогенетическим розвитком ядра консорції).
- Еволюційну.
«Таким чином, роль консорцій в системі екологічних відносин така, що вони є природними концентраторами життя в найактивніших ділянках простору (використовується з максимальною для даних умов ефективністю), що визначають спрямованість продукційних процесів і процесів споживання органічної речовини».
(Мальцев, 1987, с. 49-50).
Як подальший розвиток уявлень про взаємодію організмів між собою і середовищем існування в межах біогеоценозу виникло вчення про консорцію. Біогеоценоз – це біокосна система, яка складається із косного середовища (екотопу) та організмів, які утворюють біоценоз (Сукачев, 1964). Взаємодія між біоценозом та екотопом супроводжується кругообігом хімічних елементів і потоком енергії (Царик, Царик, 2002).
Термін консорцій уперше був запропонований ботаніком Грізенбахом (1872 – цит. за Яхонтов, 1969) для позначання спільного існування синьо-зелених водоростей з деякими трав’янистими рослинами чи водоростей та грибів, що формують лишайники.
Елементарною одиницею функціональної структури біогеоценозу є консорція. Заснов-никами вчення про консорцію стали В. М. Беклемішев (1951) та Л. Г. Раменський (1952). У своїй роботі Беклемішев відзначав, що кожен організм входить до складу БГЦ не сам по собі, а в складі якого-небудь консорція (автор застосовує даний термін у чоловічому роді). Важливо відзначити, що коли суб’єктом екологічних взаємодій, на погляд цього дослідника, є особина, то як центр консорції визначається не популяція, а особина домінуючого виду (рослини чи тварини).
В. М. Беклемішев розглядає консорцію під час обговорення тієї групи зв’язків між організмами, яку він називає прямими топічними зв’язками типу 1-а. Під прямими топічни-ми зв’язками вчений розуміє «створення однією видовою популяцією субстрату або середо-вища перебування, необхідних або хоча б сприятливих для життя іншої видової популяції». При цьому субстратом для насельника можуть служити: а) живе тіло організму – творця субстрату; б) його виділення; в) його мертві залишки; г) його спорудження. Таким чином, за В. М. Беклемішевим, консорція є сукупністю насельників живого тіла організму. Він пише: «За своїм поширенням у природі зв’язки типу 1-а – використання живого організму як субстрату для поселення іншого організму – незлічимі; огляд їх потребує цілого тому». Справді, кожний організм входить до складу біоценозу не сам по собі, а в складі якого-небудь консорцію, що складається з однієї особини виду – едифікатора консорцію й цілого ряду особин епібіонтів і ендобіонтів, що поселяються на тілі або в тілі едифікатора.
За В. М. Беклемішевим, консорція включає не популяції видів, а особини, кожний центральний вид консорції: едифікатор консорції за термінологією Беклемішева утворює в угрупованні стільки консорцій, скількома екземплярами він представлений. Консорція об-межена (за В. М. Беклемішевим) прямими топічними зв’язками з живим організмом, тому до складу консорції не входять організми, що харчуються мертвими едифікаторами, їхніми ви-діленнями й т. ін., хоча харчування органами й тканинами живого едифікатора консорції може мати місце, але не обов’язково (паразити харчуються його живими тканинами, а епіфіти – ні) (Воронов, 1974).
Е. М. Лавренком та Н. В. Дилісом (1968) був унесений ще один аспект у розуміння консорції, а саме що тільки популяція може виконувати функцію детермінанта. Завдяки пра-цям М. І. Селіванова (1974), М. А. Голубця, Ю. М. Чорнобая (1983) ці суперечності усунуто.
Сучасні погляди полягають у тому, що детермінантом консорції можуть бути як гетеротрофні, так і автотрофні організми. Консорції можна розділити на дві групи: гетеротрофно детерміновані та автотрофно детерміновані. Якщо ядром консорції є особина, то таку кон-сорцію називають індивідуальною. Індивідуальні консорції об’єднують у популяційні або інші (видові, родові, біоморфні). За походженням консорції можуть бути первинними та вто-ринними, а за складом консортів (організмів, які пов’язані з детермінантом) – повно- і не-повночленними (Царик, Царик, 2002).
Розглядаючи крім прямих і непрямих зв’язків різних типів – топічних, трофічних, фабричних і форичних, В. М. Беклемішев указує: «Розглянуті нами непрямі зв’язки являють окремий випадок більш широкого поняття – ланцюгових зв’язків». Він вважає, що дотепер щодо цього зверталася увага тільки на трофічні зв’язки – ланцюги харчування Д. Н. Кашкарова. Таким чином, ланцюгові зв’язки В. М. Беклемішев чітко відмежовує від консортивних (Воронов, 1974).
Важливою особливістю роботи В. М. Беклемішева є концепція біоценотичних зв’язків. У своїй роботі він виділив чотири типи подібного роду взаємодій: 1) трофічні (прямі й не-прямі); 2) топічні (прямі й непрямі); 3) фабричні (прямі й непрямі); форичні (прямі й не-прямі).
Л. Г. Раменський (1952) пропонує визначення консорції як «сукупність різнорідних організмів, тісно пов’язаних між собою у своїй життєдіяльності певною спільністю долі». Особливістю визначення є вказівка на спільну еволюцію (спільність долі) різнорідних орга-нізмів у складі консорції. Як приклад такої консорції автор наводить консорції деревних порід (ялина, береза, липа, дуб). На відміну від В. М. Беклемішева, Л. Г. Раменський вважав, що центром консорції може бути тільки автотрофна неепіфітна рослина. Також цей дослідник розглядав консорції під кутом взаємодій популяцій, тобто фактично заклав підвалини популяційного рівня дослідження консорцій.
Найчітше підкреслив характер міжпопуляційних відносин у своїй праці Е. М. Лавренко (1959). Він вважав консорцію сполученням популяції виду вищої рослини у даному рослинному угрупованні з пов’язаними з цією вищою рослиною популяціями нижчих рослин та тварин. Таким чином, у склад консорції цей дослідник не ввів мікроорганізми – деструктори мертвої речовини. Він також запропонував термін «консорція» замість «консорцій», який використовувався у роботі Беклемішева.
Відмінність в інтерпретації В. М. Беклемішева від поглядів Е. М. Лавренка полягає в тому, що перший автор розуміє консорцію як конкретну одиницю біоценозу (подібно поняттю фітоценозу в геоботаніці), а другий – як типологічну одиницю (подібно поняттю «асоціація» в геоботаніці).
Консорція визначається К. В. Арнольді та Л. В. Арнольді (1963) як угруповання синузіального типу, яке демонструє наявність прямих зв’язків між організмами, як результат спільного історичного розвитку та їх пристосування одне до одного. Таким чином підтримується точку зору Раменського щодо досить вузького кола організмів, які беруть участь у консорції, а також про популяційний рівень взаємодії між організмами.
Л. В. Арнольді та І. В. Борисова (1963) вважають, що тільки в межах одного біоценозу, тобто під впливом обмеженого, тривалий час більш або менш постійного набору супутніх видів можуть формуватися стійкі зв’язки центрального виду консорції та її компонентів.
Докладно розроблену методику вивчення консорції було запропоновано Л. В. Арнольді зі співавторами (Арнольди, 1960; Арнольди, Лавренко, 1960; Арнольди, Борисова, 1966; Арнольди и др., 1969). Ця методика спрямована на вивчення консортивних зв’язків комах. У процесі досліджень визначалася специфіка консортів рідкісних і звичайних рослин різних таксонів.
Якщо основою консорції є вища рослина, яка є «енергетичним пристроєм біоценозу», то така консорція вважається головною. Основою другорядної консорції є ґрунтові гриби, во-дорості, бактерії, якщо вони не є компонентами головної консорції (Арнольди, Лавренко, 1960).
Також було запропоновано основи класифікації консортів за їх просторовим розподілом та, частково, за участю у мероконсорціях окремих органів трав’янистих рослин степів Казахстану дано характеристику зв’язків консортів із центральними видами. Оскільки коло організмів у консорції з погляду цих авторів досить вузьке, то і зв’язки не відрізнялися великою різноманітні-стю. Виділялися трофічні, механічні (епіфітизм) та симбіотичні зв’язки. Другий тип є фактично топічним зв’язком із центральним видом (Арнольди и др., 1969).
За основу при вивченні консортивних зв’язків безхребетних із рослинами у працях Л. В. Арнольді використовується життєва форма. Комахи розподіляються залежно від трофічної спеціалізації, місця, яке вони займають у ярусній структурі біоценозу Екологічні особливості визначалися з урахуванням фази життєвого циклу (для личинкових та імагінальних стадій окремо). У межах ярусних екологічних груп визначаються відповідні трофічні режими членів консорцій.
У цей же час з’являються спроби класифікації консорцій за роллю їх центральних видів у формуванні рослинного покриву а також за шириною спектра живлення консортів (Емельянов, 1961, 1965). А. Ф. Ємельянов у своїх працях розрізняє такі ступені консортивних зв’язків: 1 -консортивні зв’язки монофагів; 2 - вузьких олігофагів; 3 - широких олігофагів; 4 - поліфагів. Також він розрізняє консорції домінантів, асектаторів.
Класифікація консорцій I. Л. Селіванова (1968) враховує просторове розташування консортів, а також характер взаємодії з детермінантом консорції. Автор вважав, що консортами можна назвати тільки організми, безпосередньо пов’язані з автотрофом. Ця класифікація певною мірою дає змогу охарактеризувати також і хребетних тварин, що беруть участь у роботі консорції. Важливим аспектом цієї роботи стало також визначення основних напрямків у дослідженні консорцій: флористично-фауністичного, морфолого-структурного, функціонально структурного, еколого-ценотичного, походження й еволюції консорцій.
В індивідуальній консорції відбувається елементарний акт біотичного кругообігу і перетворення енергії. Трансформации речовин та енергії в індивідуальній консорції досягаються завдяки двом процесам: синтезу речовин та енергії і деструкції мертвої органі-ки. Якщо перший процес залежить від біологічних особливостей детермінанта і діяльності консортів (паразитів, симбіонтів, фітофагів), то другий - переважно від роботи сапротрофів. Крім того, на рівні індивідуальної консорції відбувається взаємоадаптація детермінанта і консортів та зміна мікросередовища (Царик, Царик, 2002).
Ідея багатоступінчастості консортивних зв’язків була запропонована Б. А. Биковим (1967, 1970). Цей автор дійшов висновку про існування, крім консорцій автотрофних рослин, ще й консорцій з гетеротрофним ядром, які є складовими частинами більших консорцій ав-тотрофних рослин. Такі консортивні угруповання належать до консорцій другого розряду Важливим аспектом стало також описания принципів формування біогеоценозу із сукупності окремих популяцій та консорцій.
Б. А. Биковим була запропонована також класифікація консорцій. Ним виділялися такі класи:
- консорції автотрофних видів із підкласами:
■ консорції вищих рослин - домінантів рослинних угруповань суходолу;
■ великих водоростей - домінантів рослинного покриву морського шельфу;
- консорції геміавтотрофних рослин (зелені рослини, які мають елементи гетеро-трофії, наприклад, комахоїдні);
- консорції гетеротрофів (у розумінні Бикова ними є великі організми, які домінують за біомасою, чисельністю та значениям у ценозах) із підкласами:
■ консорції морських рослиноїдних тварин (в основному фітопланктонофагів) та морських хижаків;
■ наземних тварин (фітофагів та поліфагів).
У межах кожного класу можна виділити відповідні форми та різновиди консорцій: 1) ендобіонтні трофічні (з різновидами: паразитичним, симбіотичним, екрисотрофним, сапротрофним); 2) ендобіонтні топічні (медіопатичні); 3) епібіонтні трофічні (паразитичні, симбіотичні, екрисотрофні, сапротрофні); 4) епібіонтні топічні (медіопатичні).
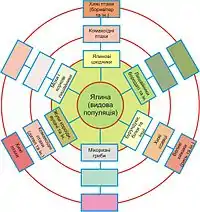
В. В. Мазінг (1966) і Т. А. Работнов (1969, 1970) вважають, що консорції являють собою ніби концентричні кола навколо центрального виду, яким є автотрофна неепіфітна вища рослина. Консорція, за В. В. Мазінгом, може бути представлена не обов’язково одним видом рослин, але групою близьких видів, наприклад сфагновими мохами. За Т. А. Работновим, до складу консорції слід включити й сапрофагів, оскільки між уживанням у їжу відмерлих частин живих рослин і мертвих, не пов’язаних із живими організмами залишків, принципової різниці немає.
В. В. Мазінг (1966) виділяє у складі консорції кілька концентрів, пропонуючи врахо-вувати у складі консорції не тільки консортів I концентра, а й організми, які впливають на I концентр (II концентр). Мазінг вважав основою консорцій трофічні зв’язки. На його думку, організми II концентра, великою мірою впливаючи на консортів I концентра, впливають на центральний вид (ядро) консорції. За аналогією ним виділялися III і IV концентри. Цей підхід був використаний О. Л. Пономаренком (2004) для візуалізації консорцій птахів у дібровах Присамар’я. При цьому Мазінг підтримував думку про те, що центральним видом (ядром) консорції може бути тільки автотрофна неепіфітна рослина, оскільки вона є елементарною «енергетичною установкою» БГЦ.
Як основу консорції Мазінг розумів не окрему особину рослини, а її популяцію. Більш широке розуміння консорції, таким чином, було сформульоване В. В. Мазінгом у визначенні: «Консорцією називається сукупність усіх організмів, пов’язаних життєдіяльністю з певним видом з автотрофних, неепіфітних вищих рослин». Також цим автором було висвітлене пи-тання про спільну еволюцію організмів, що належать до однієї консорції (Мазинг, 1970).
Консортивні відношення не можна зводити лише до ланцюга живлення. Трофічні взаємозв’язки властиві консорції, проте значна частина зв’язків є топічними, фабричними, форичними, а також медіопатичними (найменше вивчені) (Царик, Царик, 2002).
Важливий внесок у вчення про консорції зробив Т. А. Работнов (1965, 1969, 1970, 1973, 1974, 1976, 1979), який відзначав, що центром консорції можна вважати всяку автотрофну рослину (у тому числі водорості, лишайники). Він запропонував розрізняти концентри живих рослин та їх відмерлих частин. Таким чином він виділяв консортивні зв’язки консументів та редуцентів. Завдяки його працям було звернено увагу на функції мікроорганізмів, що використовують прижиттєві виділення рослин (екристотрофи), а також важливий процес розкладання мертвої органічної речовини. Також Работновим було проаналізовано характер топічних зв’язків. Він розділяв такі типи топічних зв’язків як використання тіла автотрофа як субстрату для життя та середовищетвірну роль центрального виду консорції. Він також є автором однієї з перших оглядових праць, які акумулювали усі теоретичні розро-бки того часу (Rabotnov, 1972).
А. Г. Воронов (1974) висловив такі положення, відмінні від позиції В. В. Мазінга та Т. А. Работнова:
1. Кожна особина якого-небудь виду, що є центром, ядром консорції, має безліч консортивних зв’язків. Отже, кожний екземпляр якого-небудь виду рослин, що є центральним у наведеному вище сенсі, утворює консорцію (індивідуальну консорцію за Биковим). Однак уся повнота консортивних зв’язків даного виду може бути пізнана тільки на основі вивчення всіх особин даного виду, що утворюють популяцію в рамках певного біоценозу – його ценопопуляцію (що відповідає популяційній консорції у розумінні Бикова). Таким чином, консорцію можна розглядати й у індивідуальному та у типологічному планах. Індивідуальні консорції неповні, іноді однобічні, популяційні повно відбивають характер консортивних зв’язків даного виду у фітоценозі.
2. Як основоположники вчення про консорції, так і багато авторів, що практично працюють із консорціями, відзначали з більшою або меншою чіткістю, що консорція вклю-чає тільки прямі зв’язки організмів, що належать до видів-консортів із центральним видом, який утворює ядро консорції. У цьому є глибокий зміст, тому що консорції повинні бути неоднозначні з іншими структурними, морфофункціональними одиницями біоценозу. Навряд чи є сенс створювати уявлення про консорції, якщо вони становлять явище того ж порядку, що й непрямі ланцюгові трофічні зв’язки між організмами, які лише включають, на відміну від трофічних ланцюгів, також зв’язки фабричні, форичні, топічні. Призначення поняття консорції, як вважає А. Г. Воронов, – показати, які й навколо яких організмів фокусуються комплекси прямо пов’язаних із ними й залежних від них тварин і рослин.
Екосистемний характер консорції був установлений М. А. Голубцем та Ю. М. Чорнобаєм (1983). Ці дослідники особливу увагу приділили значенню навколишнього середовища у функціонуванні консорції. Відрив консорції від зовнішнього середовища спотворює уявлення про її структуру, призводить до вилучення з її складу організмів, що пов’язані з детермінантом не лише трофічно, а й топічно, форично чи фабрично. У консорції об’єднуються всі трофічні групи організмів: автотрофи, фітофаги, зоофаги, сапрофаги, некрофаги, копрофаги, редуценти, тобто в ній відбуваються всі біотичні процеси, властиві екологічним системам, починаючи від продукування фітомаси, її споживання і переміщення через усі відомі трофічні ланцюги й закінчуючи мінералізацією.
Консорцією є така сукупність особин різноманітних видів, у функціональному центрі якої міститься особина будь-якого автотрофного чи гетеротрофного виду, компоненти якої пов’язані з цим центром трофічно, топічно, фабрично або форично, і під впливом якої формується специфічне мікросередовище (Голубець, Чорнобай, 1983).
Класифікація біогеоценозів на основі їх фізіономічної картини малоперспективна, хоча остання в окремих випадках буває й чітко позначеною. Біогеоценологічна класифікація повинна будуватися на ступені подібності процесу перетворення речовини й енергії (Сукачев, 1965), вона повинна бути типологічною, а не просторовою, – підкреслював Н. В. Тимофєєв-Ресовський (1961). При цьому не має значення склад видів, поєднуваних свідомістю людини в біоценоз; ту саму роботу виконують особини різних видів. Потрібна характеристика їх діяльності (Зубков, 1996).
У біогеоценозі яскраво виражена мозаїчність у складі як рослинного покриву, так і тваринних угруповань. На її основі можна виділити незліченну кількість екосистем різного терміну життя, розміру, але наскільки ця редукція буде відповідати критерію цілісності системи за замкненістю кругообігу речовини, настільки вона й буде органічною (Зубков, 1996). Доки таких біо(гео)ценотичних систем у літературі визнано дві: парцела (горизонтальна синузія) та консорція Беклемішева (індивідуальна консорція). Остання не володіє біогеоценотичною повнотою зв’язків, включаючи взаємодію між кожним з консортонів із собі подібними із сусідніх консорцій, тобто при роботі з розрізненими консорціями не враховується внутрішньовидова конкуренція. Це стосується в першу чергу засновника консорції – рослини-едифікатора.
Консорція, як і екоїд Негрі, не може служити базою для утворення елементарної одиниці екосистемної структури. Такою основою може стати тільки група безпосередньо взаємодіючих рослин-продуцентів. Такою групою є ценокомірка Іпатова (1966). В. І. Василевич і B. C. Іпатов (1969) поширили уявлення про ценокомірку на всі організми. З автотрофною ценокоміркою тісно пов’язані численні гетеротрофи, поєднувані взаєминами у свої ценокомірки. Сукупність косних компонентів і безпосередньо взаємодіючих між собою ценокомірок автотрофів, консументів і редуцентів, що відповідає поняттю про елементарну біогеоценотичну систему, названа «геоценоконсорцією», оскільки ці уявлення є розвитком поняття про консорцію як елементарну одиницю біогеоценозу (Зубков, 1996). Геоценоконсорції, трансгресуючи своїми рослинними ценокомірками, утворюють початковий екосистемний континуум, посилюваний перекриттям сусідніх або навіть віддалених ценоконсорцій ценокомірками рухливих гетеротрофів. Геоценоконсорція А. Ф. Зубкова (1996) близька за задумом, але не адекватна поняттям «ценекуля» Б. А. Бикова (1988) і «екоїд» Г. Негрі (Negri, 1954, цит. за Александрова, 1969). Останні два терміни-аналоги визначають ту ж саму взаємодію рослини (індивіда) з навколишнім середовищем (Зубков, 1996).
Геоценоконсорції – реальні відносно цілісні об’єкти біогеоценозу, з досить чіткими функ-ціональними зв’язками між компонентами, що існують більший час, ніж особини, що їх складають. Вони мають деяку замкнутість біоценотичних процесів, що дозволяє здійснюватися в них мінімальному біогеохімічному кругообігу речовини.
Із ценоконсорцій складаються й парцели – проміжні за ступенем цілісності субсистеми біогеоценозу. Парцели втрачають функціональну визначеність зв’язків між частинами, наявну усередині геоценоконсорції, і не набувають ще цілісності стохастичної макробіосистеми – біогеоценозу (Зубков, 1996).
Часто функціональну структуру угруповання намагаються охарактеризувати в термінах взаємодій популяцій або видів – у поняттях, запозичених з інших концептуальних рядів. Популяція являє собою репродуктивну або генетико-статистичну, а вид – таксономічну одиницю. У загальному випадку ні популяція, ні тим більше вид не обмежені рамками одного угруповання, що відразу ж робить їх використання в моделях угруповань украй незручним.
Крім того, і популяція, і вид є членами певних концептуальних ієрархій, тому їх перенесення в біоценологію призводить до появи в ній також інших понять того ж ряду (Жерихин, 1994). У випадку виду, наприклад, це таксоцени – сукупності членів одного таксона в угрупованні. Тим часом таксоцен не має функціонально-екологічного змісту, і на некоректність його використання в біоценології неодноразово вказувалося (Арнольди, Арнольди, 1963; Жерихин, 1994).
В екології виду відповідає поняття фундаментальної ніші за Д. Хатчинсоном (1965) – гіпероб’єм в n-вимірному екологічному просторі, що може бути зайнятий видом і характеризується через екологічну функцію виду й межі його толерантності. Невизначено велику кількість вимірів ніші можна для простоти звести до трьох узагальнених: організації носія ніші, використовуваних ресурсів і лімітуючих факторів, нересурсної природи (умов). Якщо обмежити діапазон ресурсів і умов рамками одного угруповання, а діапазон організації – тією частиною виду, що входить у це угруповання (ценопопуляцією), то отриманий гіпероб’єм, укладений усередині фундаментальної ніші, буде характеризувати реалізовану нішу ценопопуляції. Вона є природною й надзвичайно зручною елементарною одини-цею структури функціональних моделей (Жерихин, 1994). Ніші, поєднувані певними типами зв’язків, утворюють великі функціональні підсистеми (блоки) угруповання. Як такі блоки виступають синузії (сукупність автотрофів, що подібним чином використовують той самий ресурс (Gams, 1918)), гільдії (аналогічна сукупність гетеротрофів (Root, 1973)), консорції (вся сукупність ніш, пов’язаних прямими зв’язками з однією й тією ж нішею – ядром консорції (Мазинг, 1966)), синузіальні консорції (сукупність консорції членів однієї синузії), гільдійні консорції (те ж для членів однієї гільдії), трофічні мережі (сукупність ніш, поєднаних прямими трофічними зв’язками), трофічні рівні (сукупності ніш, що займають аналогічне положення в різних трофічних мережах), регуляторні блоки (сукупності ніш, поєднаних асиметричними за знаком зв’язками й здатних за рахунок цих зв’язків регулювати одна одну).
Кожна ніша входить одночасно в кілька функціональних блоків. Ніші в межах одного блоку можуть перекриватися, ступінь їх перекривання характеризує щільність їх упакування в блоці. У ньому можуть бути виділені також ніші домінуючих (ядерних) і підлеглих (сателітних) ценопопуляцій (Жерихин, 1994).
Література
- Дылис Н. В. О структуре консорций // Журнал общей биологии. — 1973. –№ 4. — С. 575—580.
- Корчагин А. А. Строение растительных сообществ. — Ленинград: Наука, 1976. — 320 с.
- Номоконов Л. И. Общая биогеоценология. — Ростов-на-Дону: Изд-во Ростов. ун-та, 1989. — 456 с.
- Розенберг Г. С., Мозговой Д. П., Гелашвили Д. Б. Экология. Элементы теоретических конструкций современной экологии. — Самара: СамНЦ РАН, 1999. — 396 с.
- Неронов В. М., Букварева Е. И., Бобров В. В. Зоогеография и современные задачи сохранения биоразнообразия //Успехи совр. Биол. 1993. — Т. 113, вып. 6. — С. 643—651.
- Сукачев В. Н. Основные понятия лесной биогеоценологии / В. Н. Сукачев // Основы лесной биогеоценологии. — М. : АН СССР, 1964. — 575 с.
- Жуков О. В. Екоморфічний аналіз консорцій ґрунтових тварин /О. В. Жуков // Д.: Вид–во «Свідлер А. Л.». – 2009. – 239 с. https://www.researchgate.net/publication/273318302_Ekomorficnij_analiz_konsorcij_gruntovih_tvarin
Примітки
- Мальцев В. И. Деторминирующая роль центрального вида консорции (на примере консорции манника большого) // Экология. — 1988, № 3. — С. 24-29
- Беклемишев В. Н. О классификации биоценологических (симфизиологических) связей / В. Н. Беклемишев // Бюлл. МОИП, отд. биол. — 1951. — Т. 61. — С. 3-30.
- Раменский Л. Г. О некоторых принципиальных положениях современной геоботаники / Л. Г. Раменский // Ботан. журн. — 1952. — Т. 37. — С. 181—201
- Мазинг В. П. Консорции как элементы функциональной структуры биоценозов / В. П. Мазинг // Труды МОИП. — 1966. — Т. 27. — С. 117—126.
- Мальцев В. И. Место консортивности в системе экологических отношений // Биологические науки. — 1987, № 8. — С. 46-50
Посилання
- Консорція // Словник-довідник з екології : навч.-метод. посіб. / уклад. О. Г. Лановенко, О. О. Остапішина. — Херсон : ПП Вишемирський В. С., 2013. — С. 110.
- Фабричні зв’язки // Словник-довідник з екології : навч.-метод. посіб. / уклад. О. Г. Лановенко, О. О. Остапішина. — Херсон : ПП Вишемирський В. С., 2013. — С. 181.
- Форичні зв’язки // Словник-довідник з екології : навч.-метод. посіб. / уклад. О. Г. Лановенко, О. О. Остапішина. — Херсон : ПП Вишемирський В. С., 2013. — С. 184.
- Учение о консорциях[недоступне посилання з жовтня 2019]
- Полевая геоботаника (підручник)
- Экология: Элементы теоретических конструкций
- Опыт исследования консортивной структуры островных сообществ птиц