Життя́ (Biota[1])— це явище, що є сукупністю фундаментальних загальнобіологічних ознак (метаболізму, гомеостазу, росту, розвитку, відповіді на подразнення, розмноження, еволюції тощо), які характеризують живих істот, відрізняючи їх від неживих об'єктів. Життя визначається як форма існування матерії, найхарактернішими рисами якої є обмін речовин, самооновлення та самовідтворення[2]. Вивченням властивостей життя займається наука біологія, яка покликана з'ясувати фундаментальні закони його існування та безперервності у часі.
Життя— це форма існування дієвих інструкцій з організації матерії. Якщо інструкції виконуються— тоді цей процес називається життєдіяльністю. Життю не обов'язково притаманний метаболізм, гомеостаз, ріст, розвиток, відповідь на подразнення, розмноження, еволюція на конкретному етапі існування. Заморожені організми, віруси поза клітинами, насіння до пророщення не мають цих властивостей, але обов'язково мають інструкції до виконання при сприятливих умовах.
Єдина відома на сьогодні форма життя базується на полімерах чотиривалентного карбону, де функціональну частину становлять білки, структурну та енергетичну— вуглеводи та жири, відтворювальну— нуклеїнові кислоти[5].
Живі організми разом із середовищем їх існування утворюють складні системи— екосистеми[6], які у планетарному масштабі об'єднуються у біосферу із широтною та висотною зональностями. На сьогоднішній день відомою є лише одна біосфера— земна. Передбачається можливість існування біосфер на інших планетах.
На сьогоднішній день не існує єдиноприйнятого визначення поняття Життя. Трактування поняття життя дуже різниться у природничих та математичних науках, у філософії та релігіях. Уявлення про саме життя, його походження та розвиток настільки відмінні у різних напрямках суспільної свідомості, що часто призводять до конфліктних ситуацій. Сьогодні триває полеміка між креаціоністами та еволюціоністами, суть якої полягає у суперечностях між баченнями походження, розвитку та сутності життя.
Природничі визначення
Різні природничі науки дають різні тлумачення Життя, однак, більшість сходиться на кількох фундаментальних критеріях живої матерії, які окреслюють життя як процес[7].
Життя— це функція негативної ентропії[8][9]. Життя є явищем, яке властиве відкритим або безперервним системам, що здатні самочинно знижувати власну внутрішню ентропію за рахунок речовин або вільної енергії, отриманих з довкілля і згодом виділеної у вигляді продуктів життєдіяльності. З точки зору біофізики живий об'єкт відрізняється від неживого тим, що здатен обмінюватись теплом і матерією із довкіллям. Завдяки цьому усуваються суперечності, які вникають при описі живої системи за допомогою другого закону термодинаміки— ентропія ізольованих систем повинна зростати, проте, зростання або сталість ентропії притаманне лише адіабатичним системам, які не обмінюються теплом із середовищем, а живі системи є відкритими системами.
Життя— це переважання процесів синтезу над процесами розпаду, пул енергоспоживаючих процесів зміни речовини і інших об'єктів фізичної хімії, в яких помітні два цикли (у часі): цикл регенерації необхідних речовин; цикл регенерації механізму регенерації речовини.
Життя як хімічна хвиля— багатовимірна каталітична циклічна хімічна реакція. У кожен момент часу її існування, званий часом життя, в кожному окремому ланцюзі реакції на будь-якому рівні масштабу розгляду (від молекул до класів живих організмів) присутні три матеріальні елементи: ресурс, каталізатор та результат. Елементи взаємодіють один з одним в певних тимчасових фазах. Коливанням є концентрації речовин. Кожен результат є ресурсом для наступної ланки взаємодії— хвилі концентрацій речовин.
Життя виникло, коли в процесі спонтанної хімічної ланцюгової каталітичної реакції однієї з ниток кінцевий результат виявився тотожним одному з власних ресурсів (ресурсом одного з попередніх поколінь). Всі циклічні хімічні реакції протікають без втрат інформації нескінченно довго— внаслідок чого хімічне життя генотипу вважається нескінченним. У комплексному потоці хімічних хвиль має місце ентропійне загасання, що приводить до необхідності смерті для окремих циклів хвилі (окремі молекули, клітини, організми).
Життя— особлива форма існування матерії, яка виникла на певному етапі її існування й характеризується процесами метаболізму[10]. Життя є біологічним явищем, якому притаманні: внутрішня структурованість, власний метаболізм, здатність до розмноження, спадковість та підтримка внутрішнього гомеостазу. Вивченням властивостей життя займається наука біологія, яка покликана з'ясувати фундаментальні закони його існування та безперервності у часі. Зокрема життя визначають як безперервний процес обміну матерією та енергією між організмом та довкіллям, і здатністю його (організму) до самовідтворення. Часто-густо, життя визначають як існування білкових тіл, однак, сутність виключно білкового життя була відкинута після відкриття неклітинних організмів, складених лише із єдиної молекули нуклеїнової кислоти, а також висунення Гіпотези світу РНК для пояснення виникнення самого життя.
Життя— це інформаційна структура з пам'яттю, що використовує власну внутрішню мову— систему сигналів, властивостей і методів, не пов'язана з конкретним матеріальним носієм і здатна без втрат інформації переміщуватися від одного матеріального носія до іншого, репродукуючись без втрат.
Життя біологічне— це комплексний процес, ключовою частиною якого є реакція матричного синтезу (синтез білка).
Філософські визначення
Філософські дискусії про уявлення життя беруть свої корені у античності. Платон (ліворуч) та Аристотель (праворуч) на фресці Рафаеля Санті. Аристотель вказував на пізнання за допомогою емпіричних досліджень, а Платон вказував на існування божественної сили
Бачення поняття «Життя» у філософії є дуже різним, що залежить від різних течій та напрямків цієї науки. Часто-густо, філософські концепції життя діаметрально відрізняються одна від іншої, ототожнюючи його із ідеальним існуванням, що має божественний початок; наділяючи рисами живого і неживі об'єкти; спрощуючи сутність явища життя до механічних моделей; обмежуючи рамками: органічне походить лише від життя; наділяючи усе живе розумом та здатністю до мислення тощо.
Життя у повній мірі може бути виміряне та пояснене з точки зору руху матерії та фізичних законів. Механіцисти спрощували сутність життя як явища, а здебільшого розглядали окремі живі організми як механічні конструкції.
Живими можуть вважатися лише органічні форми існування матерії, які якісно відрізняються від неживої природи. Все живе характеризується цілеспрямованістю формування своїх же життєвих сил (гомеостаз та розмноження). Віталізм у багатьох аспектах перегукується із теологічними уявленнями про життя та живу природу.
Віталізм спирається на ідею фундаментального поділу органічних і неорганічних матеріалів, і переконання, що органічний матеріал може бути отриманий лише із живих істот. Цей постулат був спростований у 1828 році, коли Фрідріх Велер синтезував сечовину з неорганічних речовин. Це так званий синтез Велера, який вважається відправною точкою сучасної органічної хімії.
Поняття життя є синтезом механіцизму та віталізму. З одного боку, життя можна описати з позицій фізичних та хімічних вимірювань, а з іншого— живі істоти мають властивості, які непритаманні неживим об'єктам. Конкретні властивості життя виникають від складності самого живого організму і закладеної у ньому програми (генетичні особливості).
Вбачає прояви життя в у неживій матерії. Античні гілозоїсти наділяли матеріальний всесвіт життям і свідомістю, населяючи їх духами та уособлюючи природу. Сучасні гілозоїсти вважають, що усі процеси у живій та неживій природі відбуваються за єдиним шаблоном, незалежно чи то будь-яка дискретна зміна, чи то підтримання тенденції. Геккель стверджував про існування єдності органічної та неорганічної природи і визначення їхніх властивостей фундаментальними законами буття. Представники аргентино-німецької нейробіологічної традиції наполягали, що будь-яка частина природи здатна поводити себе логічно та економічно, що є надрозумовою властивістю світу. Архітектор Крістофер Олександр висунув теорію Живого Всесвіту, де життя розглядається як широко розповсюджена структура, яка охоплює також і усі неживі об'єкти, зокрема будівлі.
Наділяє неживу природу не тільки рисами життя, а й свідомістю, уособлюючи її. Панпсихізм має декотрі риси схожості із анімізмом та гілозоїзмом, які лягли у його основу. Загалом панпсихісти стверджують, що Земля і Всесвіт є єдиним живим організмом, наділеним свідомістю, розумом і здатністю до мислення. Природа існує у вигляді безлічі окремих думок, які об'єднані в єдиний задум. Деякі течії панпсихізму вказують на різний ступінь розвитку свідомості у різних частин природи, відповідно не усі живі чи неживі об'єкти здатні до самоусвідомлення.
Релігійні визначення
Бог створив Всесвіт, усе живе та неживе, видиме і невидиме за шість днів в уявленні авраамітичних релігій
Народження діви Брахми та початок створення Всесвіту у розумінні дхармічних релігій
В основі практично усіх релігій закладене твердження про таїнство творення живого із неживого волею безсмертногоБога або Богів. Життя— це чудова властивість матерії, що дається і відбирається Богом. Розрізняють кінцеве (у часі) життя тіла і нескінченне життя душі. Живий організм— це такий, в тілі якого існує душа. У деяких релігіях існує уявлення реінкарнації— переселення душ або їх нескінченної мандрівки від одного індивіда до іншого, необов'язково людини. Для багатьох релігій також притаманна віра у загробне життя (рай, пекло, чистилище тощо). Більшість релігій стверджує про існування духовних істот: духів, демонів, ангелів тощо.
Відповідно до юдаїзму, християнства та ісламу життя є творінням Єдиного Бога, який є володарем життя і смерті. У Книзі Буття стверджується, що все живе було створене Богом на початку часу, між третім і шостим днями творіння. У Корані (іслам), на відміну від Біблії (християнство), немає детального опису акту творіння. Основною заповіддю божою є «не вбивай».
З-поміж основних вірувань індуїзмубуддизму, джайнізму, сикхізму та інших уособленням життя є самсара— нескінченний цикл народжень, смертей та реінкарнацій. У дхармічних релігійно-філософських системах— це уявлення про плинність усього живого, процес переходу однієї тілесної оболонки в іншу, ланцюг страждань у земному житті, кругообіг народження і смерті, трансміграція душі. У кругообізі самсари живі істоти, еволюціонують або регресують, проходять через різні форми життя: від мікробів, комах, рослин і навіть мінералів, до найвищої позиції— діви-творця Всесвіту Брахми. Становище, в якому опинилась та чи інша жива істота в ієрархії життя, залежить від якостей, набутих у минулих втіленнях, що є плодами карми, котрі істота змушена пожинати.
Відповідно до дхармічних віровчень Всесвіт є вічним та циклічним. У їхніх священних текстах описано виникнення Землі, людини та інших живих істот, котрі постійно проходять цикли створення та знищення (пралая).
Інші розуміння
Існує низка уявлень про явище життя, які заперечують усі ніші його бачення, або ж ґрунтуються на синтезі часто-густо діаметрально протилежних теорій та вчень. Окрім того термін «Життя» вживається у символічних та переносних значеннях.
Символізм
Життя часто ототожниться із біографією людини[10]. Воно є часом існування людини з моменту народження до моменту смерті. Іноді життя визначають як підсумок певних діянь та переживань окремого індивіда за будь-який проміжок часу— це є своєрідний життєпис людини, який називають Curriculum Vitae. Життя також може вживатися і для позначення існування та діяльності людського суспільства— реальна дійсність в усіх або окремих її проявах[10].
Переносні значення
У переносному значенні життям позначають різні процеси або явища, зокрема: термін або часіснування будь-чого чи будь-кого; енергію, внутрішню бадьорість, повноту духовних та моральнихсил; все, що є найдорожчим для людини, джерело радості чи щастя; існування без нужди і турбот та інші[10].
Самоусвідомлення
Собака не упізнає себе у дзеркалі, а сприймає відбиття, наче іншу тварину, оскільки не самоусвідомлює себе
Життя є властивістюіндивіда, який усвідомлює себе— «Я живу», «Я існую», «Я є», «це Я» тощо. Самоусвідомлення є важливою рисою розумного життя, оскільки усвідомлення самого себе дає суб'єктивну можливість до пізнання довколишнього світу, буття і життя, як явища, відношення себе до свого місця і ролі у загальній світобудові, а також відношення світу до себе самого. Усвідомлення себе є рефлексією самосвідомості, що спричиняється до пізнання свого внутрішнього, психологічного світу, а відповідно й до самопізнання. Усвідомлюючи себе, індивід зберігає своє «Я» в часі та просторі— узагальнює та об'єднує досвід і міркування, які були у минулому, є на даний момент часу і будуть у майбутньому[11].
Культ Живого Вогню був наявний у зороастризмі, пізніше вчення «Живого Вогню» було закладено грецькимфілософомГераклітом Ефеським (бл. 520 до н.е.-460 до н.е.). Він вважав Вогоньсимволом життя— процесу перманентного чергування народження і смерті. В уявленні Геракліта існує тотожність світового порядку, для всіх живих істот, немає Бога, а світ і його речі ніхто не створив, він існував завжди, є і буде вічно живим вогнем.
Ноосфера розглядається як окрема жива оболонка Землі наповнена розумом. В оригінальній теоріїВолодимира Вернадського, ноосфера є третім етапом, у послідовності фаз розвитку Землі, після геосфери (неживої матерії) і біосфери (біологічного життя). Так само, як виникнення життя принципово перетворило геосферу, так і поява людського пізнанняпринципово змінює біосферу. Серед складових частин ноосфери виділяють антропосферу (сукупність людей як організмів), техносферу (сукупність штучних об'єктів, створених людиною, та природних об'єктів, змінених в результаті діяльності людства) та соціосферу (сукупність соціальнихфакторів, характерних для даного етапу розвитку суспільства і його взаємодії з природою).
Ознаки живої матерії
Усі живі організми характеризуються низкою ключових параметрів, за якими їх відрізняють від неживих об'єктів. Зокрема, сюди приналежні такі критерії живої матерії:
Подразливість— це здатність живого організму переходити зі стану фізіологічного спокою у діяльний стан у відповідь на дію будь-якої сили, яку називають подразником, процес дії цієї сили— подразненням, а відповідь на нього— біологічною реакцією.
Адаптація— це індивідуальна або групова властивість живих організмів, що проявляється у вигляді реакції у поведінці організму, перебудові фізіологічних процесів або набутті нових анатомічних структур, які розвинулись за певний проміжок часу (недовгочасний або у процесі еволюції) таким чином, що підвищили виживання та репродуктивний успіх конкретного організму або виду.
Рівень організації живої матерії— це сукупність кількісних та якісних параметрів певної біологічної системи (клітина, організм, популяція іт.д.), що окреслюють умови та межі її існування. Сукупність рівнів організації утворює ієрархічну «драбину» живого. Об'єднання рівнів у ієрархії живого відбувається за принципом емерджентності— об'єднання цілісних одиниць нижчого рівня ієрархії характеризується появою нових якісних параметрів системи вищого рівня організації (клітини об'єднуються у тканини; організми одного виду утворюють популяцію тощо).
Виділяють такі основні рівні організації живої матерії:
Молекулярно-генетичний рівень
Хромосоми— приклад молекулярно-генетичної організації живого
Молекулярно-генетичний рівень живого— це сукупність інформаційно-каталітичних взаємодій біополімерів, які забезпечують збереження, обробку та передачу спадкової інформації у часі від материнського до дочірнього носіїв. Основним носієм спадкової інформації земних живих істот є ДНК, як виняток віроїди та РНК-вмісні віруси, у яких цю роль виконує РНК. Незрозумілою залишається схема передачі спадкової інформації лише у пріонів, які являють собою білок, і не містять нуклеїнових кислот.
Молекулярна організація живих систем являє собою цілу низку безперервних біохімічних циклів, які спрямовані на синтез енергії у вигляді АТФ з подальшою її витратою на реплікацію ДНК (РНК).
Клітинний рівень
Макромолекули та їх агрегації об'єднуються у клітину (схема будови евкаріотичної клітини)
Клітина є якісно новим рівнем організації живого, який об'єднує усі біохімічні та генетичні цикли в єдину систему, розмежовану із довкіллям. Ця система є базовою структурно-функціональною одиницею живої матерії (виняток— неклітинні форми), яка здатна самостійно відтворюватися.
Клітини об'єднуються у тканини (асиміляційна тканина рослин)
Клітини об'єднуються у тканини, де виконують спільні функції. Клітини у тканині втрачають індивідуальні риси, що відбувається у процесі диференціації і призводить до їх спеціалізації на виконанні якоїсь однієї або небагатьох суворо визначених функцій (нейрони— проводять електричні імпульси, міоцити— скорочуються, еритроцити— транспортують кисень тощо). Для клітин тканин властива групова поведінка, яка проявляється у однакових отриманні, обробці та відповіді на сигнали із зовнішнього середовища. Клітини тканини комунікують між собою за посередництва медіаторів і плазмодесм.
Організм— це елементарна біологічна система існування індивідуальних особин, незалежно від форми їх організації (неклітинні, одноклітинні, багатоклітинні). Одна із основних властивостей життя— розмноження,— можливе лише на організмовому рівні організації живої матерії. Життя проявляється виключно у реальному існуванні окремих організмів.
Організмовий рівень організації життя може збігатися із будь-яким із «доорганізмових» (організми вірусів існують на молекулярно-генетичному рівні; археїв— на клітинному; кишковопорожнинних— на тканинному тощо).
Генетично і морфологічно подібні організми становлять популяцію
Популяційно-видовий рівень організації живої матерії базується на поняттях популяції та виду. Популяція— це будь-яка сукупність генетично і морфологічно близьких особин, приналежних до одного виду, які заселяють визначену територію, вільно схрещуються між собою та дають здатне до розмноження потомство. Популяція є елементарною одиницею еволюції. Вид— це сукупність усіх популяцій генетично і морфологічно близьких особин, які розселені в межах певного ареалу, і між якими існують потоки генів.
Популяційно-видовий рівень організації живої матерії характеризується вільним обміном спадковою інформацією між подібними індивідуальними організмами. Це єдиний рівень взаємодії між організмами, який забезпечує вертикальний переніс генів— від батьків до дітей.
Популяційно-видовий рівень організації живої матерії забезпечує елементарні процеси еволюції, зокрема мікроеволюції.
Екосистемний рівень
Популяції різних видів об'єднуються у екосистеми. (Екосистема коралового рифу
Елементарною одиницею екосистемного рівня організації є консорція, у центрі якої знаходиться вид-детермінант, з пов'язаними із ним видами-консортами приналежними до концентрів різних порядків[21][22].
Екосистемний рівень організації живої матерії є рушієм коеволюційних процесів, ґрунтованих на міжвидових взаємодіях.
Біосферний рівень
Біосферний розподіл первинної біомаси у морських (хлорофіл А у мг/м³) та суходільних (нормована різниця індексу рослинного покриву) екосистемах впродовж 1997—1998рр. (несправжні кольори)
Біосферний рівень організації живої матерії— це сукупність усіх екосистем планети, які пов'язані між собою глобальними геохімічними циклами, океанічними та атмосферними циркуляціями.
Для цього рівня організації живого характерна агрегація однотипних екосистем, утворюючи біоми— природні зони. Біоми чітко розподілені на поверхні планети у глобальному кліматичному градієнті.
Розрізняють вісім основних суходільних біомів[23]: тундра, тайга, листопадні ліси помірного клімату, вічнозелені широколистяні субтропічні ліси, степи, пустелі, савани та тропічні дощові ліси. Окрім них, вирізняють ще шість проміжних біомів: полярні льоди, високогір'я, чапарель, вічнозелені теплі вологі ліси, тропічні мусонні ліси та напівпустелі.
В усіх клітинних організмівгенетична інформація зберігається у вигляді лінійної або кільцевої дволанцюгової антипаралельної лівозакрученої спіральної молекулиДНК. У евкаріот (за поодинокими винятками) та декотрих прокаріот ДНК з'язана із білками, переважно гістонами (евкаріоти) або гістоноподібними білками (прокаріоти), які стабілізують її структуру і захищають від гідролізації та ферментівнуклеаз. У прокаріотів ДНК являє собою одну кільцеву хромосому-плазміду.
У неклітинних організмів генетична інформація зберігається у вигляді одно- або дволанцюгових, лінійних, замкнених або кільцевих спіральних молекул ДНК чи РНК. Їх збереження відбувається багатьма різноманітними способами: від утворення комплексів із білками до вмонтовування у геном клітини-господаря.
Реалізація генетичної інформації
Схема реалізації спадкової інформації у клітині евкаріотів
Основним процесом реалізації генетичної інформації (у клітинних організмів) є власний біосинтез білка, що забезпечує структурну стабільність клітини та гомеостаз її внутрішнього середовища. Реалізація генетичної інформації включає такі стадії: транскрипцію— «перепис» інформації із матриці ДНК на матричну РНК (мРНК); процесинг— «дозрівання» матричної РНК шляхом вирізання інтронів та інших змін; трансляцію— синтез білків на мРНК.
Під час транскрипції відбувається зчитування генетичної інформації, зашифрованої в молекулах ДНК, і запис цієї інформації в молекули мРНК. Під час низки послідовних стадій процесингу з мРНК видаляються деякі фрагменти, непотрібні в подальших стадіях (сплайсинг), додавання регуляторних послідовностей (кеп, поліаденілування) і відбувається редагування нуклеотидних послідовностей. Після транспортування зрілої молекули мРНК з ядра до рибосом відбувається власне синтез білкових молекул шляхом приєднання окремих амінокислотних залишків до поліпептидного ланцюжка, що росте. На останній стадії посттрансляційної модифікації відбуваються зміни новосинтезованого білка шляхом приєднання до нього небілкових молекул та ковалентними модифікаціями його амінокислот.
У неклітинних організмів (виняток пріони) реалізація генетичної інформації відбувається шляхом використання білоксинтезуючого апарату клітини-господаря.
Також реалізацією генетичної інформації є дія некодуючих РНК — молекул РНК, які самі по собі не використовуються для синтезу білків, проте виконують інші функції, такі як захист геному від перебудови транспозонів (піРНК), регуляція активності певних генів (міРНК, мікроРНК) тощо.
Передача генетичної інформації
Передача генетичної інформації є найважливішим процесом збереження життя у часовому вимірі. Універсальним для усіх живих організмів (за винятком пріонів) є процес реплікації спадкової молекули— відтворення дочірніх молекул (ДНК або РНК) на материнській молекулі-матриці (переважно ДНК).
Реплікація ДНК
Схематичне зображення процесу реплікації. Позначення: (1) ланцюг, що відстає, (2) ланцюг-лідер, (3) ДНК-полімераза (Polα), (4) ДНК лігаза, (5) РНК-праймер, (6) ДНК-праймаза, (7) фрагмент Окадзакі, (8) ДНК-полімераза (Polδ), (9) хеліказа, (10) одиночний ланцюг зі зв'язаними білками, (11) топоізомераза
Спершу ферменттопоізомераза «розкручує» спіральну молекулу ДНК з подальшим приєднанням стабілізувальних білків, які запобігають повторній спіралізації. Фермент хеліказа розриває водневі зв'язки між азотистими основами, внаслідок чого ділянка подвійної молекули ДНК розпадається на два ланцюги. До ланцюга приєднується ДНК-праймаза— фермент який розпочинає синтез ДНК— власне реплікацію. Вона синтезує РНК-праймер— послідовність нуклеотидів від якої наступний фермент— ДНК-полімераза будує новий ланцюг, використовуючи наявний як матрицю. Синтез нових ланцюгів відбувається асиметрично, тобто один з них синтезується безперервно, а інший— будується короткими фрагментами Окадзакі у протилежному напрямку від дії хелікази. Фрагменти Оказакі з'єднує між собою фермент ДНК-лігаза. Таким чином з однієї молекули ДНК утворюються дві ідентичні, які після закінчення процесу реплікації спіралізуються.
Реплікація у вірусів що мають одноланцюгову ДНК має особливості. У клітині хазяїна на такій молекулі, яку називають (+)-ланцюгом синтезується комплементарний йому (-)-ланцюг, таким чином утворюється дволанцюгова молекула ДНК. (-)-ланцюг потім слугує матрицею для синтезу нових (+)-ланцюгів, які вбудовуються у вірусні частинки. У процесі беруть участь ферменти вірусів та ферменти клітини-хазяїна.
Реплікація РНК
Реплікація РНК відбувається у організмів, геном яких кодує ця нуклеїнова кислота— це деякі типи вірусів та віроїди. Процес відбувається в клітинах хазяїна, які були інфіковані цими організмами. При цьому також синтезуються (-)-ланцюги та РНК проходить дволанцюгову стадію.
Процес онтогенезу є безперервним і пов'язаний із послідовною активацією та дезактивацією груп генів, які відповідають за розвиток організму у конкретний період його життя. У молодих організмів процеси анаболізму (синтезу) переважають над процесами катаболізму (розпаду); у зрілих організмів, які активно розмножуються, зберігається рівновага між обома проявами метаболізму; організми, що старіють, характеризуються переважанням катаболізму.
Наука, яка вивчає онтогенез називається Біологією індивідуального розвитку (БІР).
Між онтогенезами тварин, рослин і грибів є суттєві відмінності. Найбільш типово індивідуальний розвиток вивчений на тваринах, у яких він є найскладнішим з-поміж усіх живих організмів на Землі.
Онтогенез тварин
Паралелізм у ембріональному розвитку усіх класівхребетних
Онтогенез тварин включає такі стадії:
1. Ембріональна— зародкова стадія, що триває з моменту утворення зиготи до народження або виходу з яєчних оболонок. Його основними етапами є: морула, бластула, гаструла, зародок, зростання і народження особини. Після запліднення починається дробіння яйця, яке має характер палінтомічного поділу. Дробіння завершується утворенням одношарового зародка— бластули. Після закінчення дробіння клітина ділиться монотомічно, починаються процеси диференціації та переміщення клітин (гаструляція), які спочатку приводять до утворення двох або трьох зародкових листків (ектодерми, ентодерми та мезодерми), а пізніше— зачатків органів. Від цієї схеми відрізняється онтогенез первинних багатоклітинних (Prometazoa), які не мають зародкових листків.
2. Постембріональна— післязародкова стадія наступає після виходу молодого організму з яєчних оболонок або народження, і триває до набуття організмом здатності до розмноження. Після виходу з яйця (або народження) починається постембріональний розвиток, який буває прямим (коли молодий організм подібний до дорослого) або супроводжується метаморфозом.
3. Статева зрілість— ця стадія наступає після досягнення молодим організмом дорослого стану, коли він готовий до статевого розмноження.
4. Старіння— стадія розпочинається з часу втрати організмом здатності до розмноження і до моменту його смерті.
Онтогенез рослин
Схема онтогенезу квіткових рослин
Основні стадії онтогенезу рослин:
1. Іматурна— зародкова стадія у рослин, що триває від моменту утворення зиготи до формування діаспори та її проростання. У різних відділів рослин іматурна стадія розвивається по-різному. Для вищих спорових у ембріогенезі притаманне формування спори, яка проростає у гаметофіт, де відбувається статеве розмноження і проростання ювенільної рослини. Для голонасінних властиве просте запліднення спермієм відкритого (голого) насінного зачатку. Для покритонасінних характерне подвійне запліднення— два спермії запліднюють дві клітини у зав'язі: гаплоїднуяйцеклітину та диплоїдну клітину зародкового мішечка. Після запліднення з насінних зачатків формується насіння. Насіння має зародок з корінцем, бруньками і сім'ядолями, запас поживних речовин, та вкрите насінною шкіркою.
2. Ювенільна («дитяча»)— від проростання насінини до формування основних органів (за винятком органів розмноження) і тканин.
3. Віргінільна («дівоча»)— стадія вегетативного росту і досягнення репродуктивного віку.
4. Генеративна— стадія активного розмноження.
5. Сенільна— від втрати здатності до розмноження і до смерті.
Онтогенез грибів
Індивідуальний розвиток у грибів немає певних універсальних етапів, що пов'язано із великою різноманітністю їхніх життєвих циклів. У загальних рисах виділяють чотири основні стадії онтогенезу грибів:
1. Зиготична стадія— наступає після статевого процесу. У більшості грибів утворюється не зигота, а клітина злиття з дикаріонтичнимиядрами. Зигота у класичному розумінні цього терміну в онтогенезі може бути взагалі відсутня, хоча замість неї після каріогамії обов'язково утворюються диплоїдні зиготичні ядра. Такі ядра є гомологами зиготи, і при розрахунку схеми життєвого циклу саме їх визначають як зиготу.
2. Дикаріонтична стадія— наявність у міцелії (незалежно чи міцелій септований, чи несептований) попарно зближених, генетично різних, гаплоїдних ядер. Для грибів у більшості випадків характерні життєві цикли з наявністю дикаріонтичних поколінь, і лише у частини хітридіомікотових та зигомікотових грибів такі покоління відсутні.
3. Анаморфна стадія— нестатеве покоління, яке здатне до розмноження за допомогою зооспор, спорангієспор чи конідій.
4. Теломорфна стадія— статеве покоління, яке розмножується за допомогою гамет, недиференційованих гаметангіїв, статевих спор (аскоспори чи базидіоспори). Вищі гриби на цій стадії утворюють плодові тіла, які несуть на собі органи статевого спороношення (аски або базидії).
Онтогенез одноклітинних організмів
Онтогенез усіх евкаріотичниходноклітинних організмів (амеби, евглени, інфузорії та ін) збігається із клітинним циклом, і представлений такими стадіями:
1. Інтерфаза— період клітинного росту, синтез мРНК та білків. Наприкінці інтерфази іде підготовка до поділу клітини, відбувається реплікація ДНК. Інтерфаза складається із таких періодів:
* G1-періоду (від англ. gap— проміжок)— період початкового росту, коли у клітині активно відбуваються процеси транскрипції і трансляції.
* S-періоду (від англ. synthesis— синтез)— період протікання реплікації та подвоєння центріолей, а за їх відсутності— базальних тілець джгутиків.
* G2-періоду— готування до мітозу.
2. Мітоз (від давн. грец. μίτος— нитка), часто каріокінез— поділ ядраеукаріотичноїклітини із збереженням числа хромосом. Мітоз складається із чотирьох етапів: профази, метафази, анафази і телофази.
* Профаза— відбувається конденсація хромосом та поділ клітинного центру.
* Метафаза— утворення метафазної пластинки.
* Анафаза— хромосоми діляться і розходяться до полюсів ділення.
* Телофаза— руйнування веретена поділу і утворення ядерної оболонки навколо дочірніх ядер.
3. Цитокінез— поділ материнської евкаріотичноїклітини навпіл із рівним розподілом клітинних органел.
Онтогенез неклітинних організмів
Онтогенез неклітинних організмів вирізняється з-поміж інших живих істот ключовим процесом реплікаціїнуклеїнової кислоти. Єдиним винятком є аміноаквеї— пріони, у яких нуклеїнові кислоти відсутні. В залежності від складності неклітинних форм життя їх онтогенез включає такі стадії:
1. Реплікація— процес відтворення ДНК/РНК. Характерний для усіх нуклеаквеїв, проте, відсутній у аміноаквеїв.
2. Транскрипція— процес синтезу мРНК на матриці ДНК/РНК. Характерний лише для вірусів.
3. Трансляція— процес синтезу білків на мРНК. Характерний лише для вірусів.
4. Збір віріонів— процес пакування спадкової молекули нуклеїнової кислоти у білковийкапсид. Характерний лише для вірусів.
Філогенез
Філогенетичне дерево Хребетних: рептилії та птахи є монофілетичною гілкою еволюції; різні групи рептилій є парафілетичними одна відносно іншої; птахи і ссавці є поліфілетичними групами хребетних
Філогенез— історичний розвиток як окремих видів і систематичних груп організмів, так і органічного світу в цілому. Філогенез взаємозв'язаний з онтогенезом.
Філогенез— послідовність подій еволюційного розвитку виду або таксономічної групи організмів. Вивчає філогенез і класифікує організми на його основі філогенетика.
Виділяють такі форми філогенезу:
1. Дивергенція— розходження ознак і властивостей у спочатку близьких груп організмів у ході еволюції. Вони виникають в результаті різнонапрямленого природного добору, який виникає під дією відмінних одних від інших умов зовнішнього середовища. Під їхньою дією формується адаптивна радіація ознак живих організмів— тип еволюції, при якому дві або більше біологічні характеристики мають спільне походження, але розійшлися впродовж еволюційного процесу. Ці характеристики можуть бути як морфологічними рисами, так і молекулярними, наприклад генами, сигнальними або метаболічними шляхами.
2. Конвергенція— це еволюційний процес, результатом якого є утворення схожих морфологічних ознак у представників неспоріднених (немонофілетичних) груп (наприклад, хвостові плавці риб, вимерлихрептилійіхтіозаврів та ссавцівкитів). Основною причиною конвергенції є однакові умови середовища існування живих організмів (наприклад, риби, іхтіозаври та кити— мешканці водного середовища).
3. Паралелізм— незалежне виникнення схожих морфологічних чи фізіологічних ознак у близькоспоріднених груп організмів (наприклад, у різних груп динозаврів розвинулись ознаки птахів: пір'я, дзьоб, політ тощо), які розвинулись внаслідок існування у однакових умовах середовища та однонапрямного природного добору.
Класифікація живих організмів
Ієрархія біологічної систематики
Класифікацією живих організмів називають стандартизоване групування істот (живих і вимерлих) за принципами еволюційних зв'язків на базі морфо-анатомічної, ембріологічної та молекулярної подібностей, об'єднуючи їх у ієрархічнусистему, в основі якої лежить поняття біологічного виду. Систематична ієрархія об'єднує еволюційно близькі види у роди, роди— у родини, а їх у ряди (порядки), далі у класи, типи (відділи), царства й домени.
Сучасна систематика, побудована на основі молекулярної (ДНК, РНК) спорідненості між живими організмами, перебуває на стадії свого становлення, переживає революційні зміни, тому нині існує кілька альтернативних систем класифікації живих організмів. Це особливо стосується вищих таксонів, таких як домени, царства і типи; у меншій мірі класів та рядів. З-поміж систематиків не існує одностайності щодо бачення загальноприйнятної класифікації життя на Землі. Найбільш кардинальні систематичні зміни торкнулись найпростіших, яких розподілили не лише між окремими філами, а й царствами[25].
Історичні зміни уявлень про вищі таксони живих істот
Все ще ведуться дискусії щодо доцільності виділення філ дво- і одноджгутикових[27].
Форми існування життя
Усі живі форми на Землі поділяються на клітинні та неклітинніорганізми. Базовою одиницею існування клітинних організмів є клітина, яка може мати різний рівень організації та складності будови. До клітинних організмів належать усі, без винятку, бактерії, археї— об'єднані під назвою прокаріоти, а також еукаріоти (одноклітинні, водорості, рослини, гриби і тварини), які можуть існувати у формі одноклітинних, тканинних чи системно-органних організмів. До неклітинних організмів належать істоти, які проявляють ознаки живого лише у внутрішньому середовищі клітинних організмів, а поза ним характеризуються ознаками неживих об'єктів. Плани будови неклітинних організмів є дуже різноманітними і базуються на молекулахнуклеїнових кислот або білків. До неклітинних організмів зачисляють: віруси, віроїди, сателіти, косміди, фазміди, плазміди, транспозони[28] та пріони[29]. Більшість із них (за винятком вірусів) часто не розпізнає як живі організми, а лише як автономні генетичні і структурні частини клітин, проте, які можуть самостійно відтворюватись і часто мають патогенну і трансмісивну здатності.
Неклітинне життя
Усі неклітинні живі організми знаходяться на молекулярному рівні організації. Вони являють собою впорядковані структури біополімерів, які несуть власну генетичну інформацію, необхідну для їх подальшого відтворення. Винятком є лише пріони, які не містять у своїй структурі ДНК— носія генетичної інформації.
Нуклеїновмісні організми або нуклеаквеї
Розфарбована сканувальна мікроелектронна світлина вірусу грипу (Influenzavirus)— приклад найскладнішої молекулярної форми життяВіроїд серцевини бульб картоплі (PSTV)— схема вторинної структури одноланцюгової замкненої РНКВіруси (Vira)— облігатні внутрішньоклітинні паразитиклітинних організмів, представлені нуклеїновою кислотою (ДНК або РНК) і білковою оболонкою. Розміри коливаються від 10 до 300 нм. Розмножуючись тільки в живих клітинах, вони використовують їхній ферментивний апарат і переключають клітину на синтез зрілих вірусних часток— віріонів. Поширені всюди. Викликають захворювання бактерій, архей та евкаріотів.
Віроїди (Viroides)— облігатні внутрішньоклітинні паразитирослин, представлені висококомплементарною одноланцюговою кільцевою молекулою РНК, розміром 250—375 нуклеотидів[30]. Віроїдоподібні організми виявлені також і у тварин— Гепатитний дельта вірус (HDV), асоційований із Вірусом гепатиту В (Orthohepadnavirus HBV)[30]. Близькими до віроїдів є також ретровіроїди— незалежні, неінфекційні РНК—коменсали рослин, які виступають матрицею для синтезу ДНК, вмонтовуючись у геномклітини-господаря[30]. РНК віроїдів не кодує жодного білка, однак, конкурує за доступність клітинногоферментуРНК-полімерази. Походження віроїдів залишається невідомим. Існують гіпотези про їх виникнення від інтронів або транспозонів. Припускається, що віроїди можуть виникати при рекомбінаціях клітинної ДНК.
Сателіти (Satellites)— облігатні внутрішньоклітинні паразитирослин, представлені лінійною РНК завдовжки 332—405 нуклеотидів[31]. Сателіти асоційовані із Вірусом огіркової мозаїки (Cucumovirus), з яким передаються від рослини до рослини. Відомий вид CARNA-5 спричиняє загибельпомідорів та деяких інших рослин[32]. У 1977 році у Франції, Італії та Іспанії спалахнула епідемія CARNA-5, що призвела до цілковитої загибелі помідорних плантацій.
Косміди (Cosmides)— облігатні внутрішньоклітинні паразити і мутуалісти різних клітинних живих організмів, представлені великими дволанцюговими кільцевими ДНКплазмідами, завбільшки 37-52 тисячі пар нуклеотидів. Косміди асоційовані із Лямбда бактеріофагами, здатні проникати у його капсид під час запаковування фаговоїДНК, що не є властивим для плазмід. Таким чином косміди передаються від одного організму до іншого.
Фазміди (Phasmides)— облігатні внутрішньоклітинні мутуалісти та коменсалибактерій, які являють собою одноланцюгові кільцеві ДНКплазміди. Фазміди асоційовані із багатьма бактеріофагами, з якими гібридизуються. Вони здатні вмонтовуватись у геном бактерії-господаря, реплікуватися разом з ним, захоплювати його окремі гени і запаковуватись у капсидбактеріофагів, з якими передаються до інших бактерій. Фазміди часто називають ще фаґемідами— клонувальним вектором, використовуючи у генній інженерії для переносу генів.
Кільцева дволанцюгова ДНК плазміда під растровим електронним мікроскопомПлазміди (Plasmides)— облігатні внутрішньоклітинні мутуалісти, коменсали або паразити, представлені кільцевою дволанцюговою ДНК, розміром від 1тис. до 1млн пар нуклеотидів. Вченідискутують щодо визнання чи не визнання плазмід самостійними організмами. Одні автори сходяться на тому, що плазміди не є окремими організмами[33], тоді як відповідно до декотрих класифікацій, їх розглядають як повноцінні живі організми на молекулярному рівні організації[34].
Плазміди пройшли довгий еволюційний шлях разом зі своїми господарями, тому вважаються мобільною частиною їх геному— мобіломи,— однак, реплікуються цілком незалежно від нього[35], оскільки є самостійними репліконами. Вони найбільш притаманні для бактерій, проте, виявлені також у археїв та евкаріотів.
Плазміди задіяні у горизонтальному переносі генів між різними бактеріальними клітинами.
Розрізняють п'ять великих груп плазмід: статеві (F-плазміди), резистентності (R-плазміди), коліцинові (Col-плазміди), травні та вірулентні.
Плазміди, які існують у батьківській клітині в одному або лише в кількох екземплярах, виробили шляхи активного розповсюдження в дочірніх клітинах. Один із них— це одночасне продукування білковоїотрути (має тривалий термін дії) та антидоту до неї (діє дуже короткий час). Якщо при поділі материнської у дочірню клітину не потрапить копія плазміди, то дія протиотрути у ній припиняється і клітина гине від токсичного шоку[36].
Схема будови бактеріального композитного ДНК-транспозонуТранспозони— облігатні внутрішньогеномні паразити[37], близькі до вірусів, що здатні змінювати своє положення у геноміхазяїна. Транспозони ще називають егоїстичною ДНК, оскільки вони здатні самовідтворюватись і розповсюджуватись по геному, незалежно від останнього, а також не вносять (за невеликими винятками) вкладу у репродуктивну здатністьорганізму. Структурно, транспозони являють собою послідовності ДНК, що можуть переміщатися в межах геному однієї клітини за допомогою процесу транспозиції. В результаті цього процесу вони можуть викликати мутації і змінювати кількість ДНК в геномі. Транспозони також відомі під назвою «гени, що стрибають» і є прикладами мобільних генетичних елементів. Існує два типи транспозонів: І та ІІ класів. Транспозони І класу— ретротранспозони— нагадують ретровіруси, оскільки переміщуються по геному шляхом транскрипції з ДНК у РНК, а потім назад до ДНК за допомогою зворотної транскриптази. Транспозони ІІ класу переміщуються по геному уникаючи РНК-стадії, а «випадаючи» і «вмонтовуючись» у вигляді ланцюга ДНК. Виникнення та еволюція транспозонів досі залишаються незрозумілими, вважається, що вони мають спільного предка із декотрими родинами вірусів.
Безнуклеїнові білкові організми або аміноаквеї
Просторова структура коров'ячого пріону PrPCПріони— внутрішньоклітинні паразитинейронівссавцівбілковоїприроди. Пріоноподібні білки виявлені у деяких грибів, однак, вони не спричинюють захворювань своїх господарів[38].
Пріони являють собою просторово видозмінені білки, які перейшли із α-спіралі до β-шару, що надає їм надзвичайної стійкості до фізичних, хімічних та біологічних факторів денатурації білків. Пріони нездатні до розмноження, однак відтворюються шляхом каталізу просторової перебудови нормальних гомологічних їм білків нейронів, спричиняючи ланцюгову реакцію перетворення білків і розвиток захворювання[39]. Пріони потрапляють в організм через травну систему із їжею, проникають у кров і мігрують до мозку.
Пріоноподібні білки (PrP) містяться на мембранах більшості клітин організму, складаючись із 209-ти амінокислот. Нормальні пріоноподібні білки позначають PrPC, де «С» означає «клітинний» або «звичайний». Їх функція до кінця не з'ясована, відомо, що PrPC мають високу спорідненість до іонівкупруму[40]. За невідомих обставин PrPC можуть переходити із α-спіралі до β-шару, утворюючи інфекційні білки, які позначають PrPSc. «Sc» означає «Скрепі»— назва походить від пріонного захворювання овець, що називається «овеча сверблячка» або по-англійськи «скрепі». PrPSc здатні «склеюватись» із нормальними PrPC білками, порушуючи фізіологічний стан окремих нейронів та руйнуючи цілісність мозку.
Пріони викликають низку смертельних захворюваньссавців: куру та хворобу Кройцфельда-Якоба у людей, коров'ячу губчасту енцефалопатію (коров'ячий сказ) у корів, овечу сверблячку (скрепі) у овець тощо.
Основна характеристика одноклітинної форми існування живих організмів полягає в тому, що окрема індивідуальна особина, представлена лише єдиною клітиною, поєднує в собі всі функції, притаманні й для багатоклітинних організмів. Для одноклітинних організмів характерні всі фундаментальні властивості живого. Одноклітинними формами представлені усі прокаріоти (бактерії, археї) та деякі евкаріоти (інфузорії, динофлагеляти, мікроспоридії, деякі тварини (комірцеві джугтиконосці), декотрі рослини (вольвокс) та ін.).
Гелікобактер Пілора (Helicobacter pylori)— одноклітинний прокаріотичний організмПрокаріоти— це збірна група одноклітинних або колоніальних живих організмів, клітини яких позбавлені морфологічно сформованого (відокремленого каріоплазмалемою) ядра, а також різноманітних мембраннихорганел (винятком є синьо-зелені водорості). Прокаріоти мають типову клітинну будову, що передбачає відмежування їх внутрішнього середовища (цитоплазми) від зовнішнього клітинною мембраною, а у більшості наявні ще й клітинні стінки та слизові капсули. Розміри клітин прокаріотів коливаються в межах 0,5-2 мкм, винятком є лише нанобактерії, розміри яких є меншими за загальноприйняту нижню межу розмірів клітини, і становлять від 30 до 200 нм.
Характерною особливістю клітинної будови прокаріотів є відсутність клітинних компартментів, утворених мембранами, розвиненого цитоскелету, ендосимбіотичних органел (мітохондрії, пластиди) та організованої у хромосомиДНК. Їхня ДНК замкнена у кільце і нагадує плазміду, а геном організований в оперони. Білоксинтетичний апарат представлений, так званими, «малими рибосомами»: велика субодиниця масою 50S (сведберґів), мала субодиниця— 30S. Деякі прокаріоти мають внутрішньоклітинні мембранні утвори— ламели, на яких містяться комплекси цитохромних білків: фотосинтетичний апарат або дихальний ланцюг.
До прокаріотів відносять представників двох доменів: бактерії та археїХаос каролінський (Chaos carolinense)— одноклітинний евкаріотичний організмЕвкаріотичнінайпростіші— це збірна група різних за походженням та морфологією одноклітинних або колоніальних живих організмів, у клітинах яких наявне ядро і комплекс різноманітних мембранних органел (ЕПС, апарат Ґольджі, пластиди, мітохондрії тощо)[41]. До одноклітинних традиційно відносять амеб, інфузорій, слизовиків, евґлен, апікомплексних, джгутикових, опалін, лабіринтових та багатьох інших.
За планами клітинної будови найпростіші є дуже різноманітними. Клітинні покриви представлені як плазмалемою (амеби, слизовики), так і пелікулярними (евґленові), целюлозо-пектиновими (жовтозелені водорості) й альвеольованими (інфузорії, динофлаґеляти) клітинними оболонками тощо. Більшість із них має гетеротрофний (тваринний) спосіб живлення, для вторинних фотосинтетиків притаманна міксотрофія, і тільки для незначного числа найпростіших властива автотрофність. Поглинання їжі відбувається шляхом фагоцитозу або піноцитозу, а у декотрих груп (інфузорії) існують спеціальні пристосування для вловлення і поглинання їжі. Їжа перетравлюється у травних вакуолях — це називається клітинним травленням.
Одноклітинна інфузорія Трубач Резела (Stentor roeseli). На малюнку праворуч помітне вегетативне ядро— макронуклеус.
У клітинах найпростіших наявне одне, два і більше ядер. У декотрих наявні диплоїдне генеративне та вегетативне ядра (інфузорії), у інших— лише гаплоїдні ядра (апікомплексні), або лише диплоїдні (евґленові). Найпростіші мають типову для евкаріот хромосомну організацію ядерної лінійної ДНК зв'язану із білкамигістонами. Їх клітини поділені на компартменти складною системою мембран. Добре розвинений цитоскелет утворений тубуліновимимікротрубочками та актиновимимікрофіламентами, які надають форми клітині та дозволяють змінювати її форму. Також розвинений джгутиковий апарат, який забезпечує переміщення більшості найпростіших у просторі. Центром веретена поділу виступають центріолі або базальні тіла джгутиків. У життєвому циклі усіх найпростіших наявний простий— мітоз, а у деяких також редукційний поділи клітини— мейоз. Життєві цикли різноманітних планів із чергуванням поколінь і переважанням певної ядерної фази (диплофазні, дипло-гаплофазні, гапло-диплофазні) або цикломорфозу. Для усіх найпростіших (за винятком «енергетичних паразитів» мікроспоридії, метамонади) притаманні первинні ендосимбіотичні органели мітохондрії, а для багатьох— вторинні ендосимбіонти хлоропласти хлорофітного або родофітного (походять або від зелених або від червоних водоростей) типів.
Різноманіття форм одноклітинних діатомових водоростей
За будовою мітохондрій, як найдревніших первинних ендосимбіонтів, найпростіші (багатоклітинні також) відносяться до кількох різних груп (царств)[42]:
Гіпохондріати або первинно безмітохондріальні— реліктові внутрішньоклітинні («енергетичні») паразити позбавлені мітохондрій. Їх ще називають домітохондріальними. Сюди належать Мікроспоридії та метамонади.
Дискокристати— організми, у клітинах яких містять найпримітивніші мітохондрії із трьома типами крист (складки внутрішньої мембрани мітохондрії): дископодібними, трубчастими і пластинчастими. Дискокристати є базовою групою диверґенції інших евкаріот. Сюди приналежні евґленові, кінетопластиди, ахразіомікоти та ін.
Тубулокристати— організми з двома типами мітохондріальних крист: трубчастим та пластинчастим. Група об'єднує як одноклітинні (амеби, слизовики, інфузорії, діатомові водорості), так і багатоклітинні (бурі водорості) форми.
Платикристати— організми, у клітинах яких наявні мітохондрії виключно із пластинчастими кристами. Сюди приналежні багатоклітинні організми і лише незначна частка видів має одноклітинну життєву форму (комірцеві джгутикові, вольвоксові, хітридіомікотові та деякі інші).
Нитчасті та несправжньотканинні організми
Нитчастий план будови водорості спіроґіри (Spirogyra)Плодове тіло їстівного білого гриба (Boletus edulis) утворене несправжньою тканиною— плектенхімоюГубки— дотканинні тварини
Нитчастий та несправжньоклітинний плани будови багатоклітинних організмів притаманні для декотрих водоростей, більшості вищих грибів та нетканинних тварин: пластинчастих, губок, ортонектид та дицимиїд.
Нитчасті форми будови тіла характеризуються недиференційованими, однак фізіологічно та функціонально пов'язаними між собою клітинами, які розташовані одна над іншою, утворюючи нитку (трихому). Наростання нитки відбувається виключно за рахунок поділу верхівкової клітини. Морфологічно та функціонально відмінними є лише верхівкова— виконує функцію наростання; та базальна клітини— виконує функцію кріплення до субстрату.
Розрізняють прості трихальні та розгалужені— гетеротрихальні таломи притаманні для водоростей (деякі зелені та червоні водорості). Деякі одноклітинні водорості (діатомові) можуть утворювати несправжні нитки, клітини у яких не пов'язані одна з іншою. У грибів нитчаста будова вегетативного тіла утворена септованим міцелієм, який поділений на клітини поперечними перегородками— септами (аскомікотові та базидіомікотові гриби).
Модифікацією нитчастої структури тіла є злиття клітин у єдину велетенську багатоядерну структуру, яка у водоростей називається сифональним або сифонокладальним (неповне злиття) таломом (жовто-зелені водорості), а у грибів— несептованим міцелієм (оомікотові гриби, зигомікотові гриби).
Трихоми здатні зростатися (сплітатися) між собою та формувати несправжню тканину. Така несправжня тканина формує таломи багатоклітинних зелених і червоних водоростей, а також плодові тілагрибів і слань лишайників. Її називають псевдопаренхімою у водоростей та плектенхімою у грибів. У грибів плектенхіма часто диференціюється і утворює покривні структури, однак зберігає свою нитчасту будову. Особливістю несправжніх тканин є їх нездатність до поділу в різних площинах, а лише в одній.
Несправжні тканини також утворюють деякі синьо-зелені водорості (стигонематальні), їх клітини можуть поділятися у різних площинах, утворюючи паренхіматозні структури. Однак, на відміну від справжніх тканин, клітини стигонематальних функціонально недиференційовані.
Дотканинна будова також характерна для тварин, приналежних до типугубки, тіло яких має поклітинну диференціацію, коли будь-яка клітина тіла може у певний момент часу змінити свою функціональну приналежність і замінити будь-яку іншу клітину. Їх зовнішня та внутрішня будова настільки незвичайні, що тривалий час учені не знали, до якого царства живих істот вони належать, і лише в 1825р. їх було визначено як тварин[43]. Тіло губок побудоване з двох шарів клітин: пінакодерми, що вкриває тіло ззовні, та хоанодерми, яка вистилає парагастральну порожнину або джгутикові камери. Між цими двома шарами розташований мезохіл (паренхіма, мезенхіма), який складається з клітин різноманітної будови та функцій (археоцити, коленцити, лофоцити, склероцити та ін.) і продуктів їх виділення, він містить також скелетні елементи. Товща мезохілу пронизана каналами, що відкриваються на поверхні невеличкими отворами— порами.
Тканинні організми
Бура водорість макроцист вогнеплідний (Macrocystis pyrifera) — приклад тканинних організмівАурелія вухата (Aurelia aurita)— організм із тканинною організацією, позбавлений органів
Тканинні організми є багатоклітинними живими істотами, клітини яких згруповані у тканини із певною функціональною спеціалізацією (нервова, травна, механічна, епітеліальна тощо), однак вони не утворюють органів.
Прикладом тканинної організації є бурі водорості та деякі тварини (кишковопорожнинні та реброплави).
Для декотрих порядківбурих водоростей характерні таломи тканинної будови. У найпростішому випадку такий талом складається лише з двох типів тканин— кори та серцевини. Кора утворена дрібними забарвленими клітинами, що виконують функцію фотосинтезу. З них також утворюються органи розмноження. Серцевина складається з великих, переважно безбарвних клітин, що виконують провідну та запасальну функції.
У складніших випадках талом утворений більшою кількістю тканин: меристодермою, корою, проміжною тканиною та серцевиною. Меристодерма— це поверхнева тканина, що виконує захисну та репродуктивну функції. Клітини меристодерми звичайно дрібні і здатні до активного поділу. Кора утворена більшими за розміром клітинами з добре розвиненими хлоропластами. Основною функцією кори є здійснення процесу фотосинтезу. У клітинах проміжної тканини накопичуються продукти асиміляції. Самі клітини великі, звичайно з редукованими хлоропластами. Серцевина утворена ситовидними трубками та трубчастими нитками.
Двошарові тварини
Для кишковопорожнинних характерний тканинний тип організації, тобто клітини в їх тілі об'єднані в тканини. Проте вони не мають складних органів, властивих більш високоорганізованим тваринам. Важливою прогресивною рисою кишковопорожнинних є поява в них нервової системи та м'язових (епітеліально-м'язових) клітин. За рахунок цього тварини сприймають різні подразнення та рухаються. Кишковопорожнинні— більш інтегровані тварини порівняно з Prometazoa— у них з'являються загальноорганізмові функції— живлення та рух. Живляться вони активно, а не пасивно, захоплюючи та вбиваючи здобич; переважна більшість їх— хижаки. Хижий спосіб життя сприяє інтеграції організму й подальшій еволюції.
Частина «стебла» моху гілокому блискучого (Hylocomium splendens)— приклад примітивної диференціації на органи у рослинЧастина вегетативного тіла кропиви дводомної (Urtica dioica: видно диференціацію на стебло, листки та квітки (суцвіття)
Зелені рослини є системно-органними живими організмами, яким притаманна диференціація вегетативного тіла на органи: стебло, корінь (ризоїд), листок, квітку (стробілу), які об'єднані в єдине провідною системою. У різних відділів розвиток тих чи інших органів є відмінним.
Несудинні рослини
До несудинних належать мохи та печіночники.
Основною характеристикою несудинних рослин є відсутність провідної системи, зокрема трахеїд та судин. Тіло представлене гаметофітом і диференційоване на несправжнє стебельце (каулідій) з дрібними несправжніми листками-філідіями без справжніх коренів. Окрім статевого розмноження гаметофіт мохоподібних виконує функції живлення, водопостачання та фотосинтезу. На гаплоїдному гаметофіті розвиваються чоловічі (антеридії) та жіночі (архегонії) статеві органи. Диплоїднийспорофіт (спорогон) дуже редукований, найчастіше паразитує на гаметофіті. Він побудований із спороносної коробочки і ніжки з гаусторієм, яким присмоктується до гаметофіта.
Судинні рослини
Нереїда бурштинова (Nereis succinea) належить до первинноротих організмівЕмбріологічний розвиток первинноротих характеризуються різноплановістю дробіння яйцеклітини: спіральне, білатеральне, радіальне та поверхневе, як правило, цей процес є детермінованим. У результаті дробіння утворюється бластула із отвором— бластопором, яка шляхом різних способів (уні- та мультиполярна імміграція, деламінація, епіболія) перетворюється на двошарову гаструлу. Бластопор гаструли перетворюється на ротовий отвір, а на протилежному або «черевному» боці проривається анальний отвір. При щілиноподібному бластопорі (круглі черви) він заростає по всій довжині, ззаду наперед. Передній залишок бластопора перетворюється на ротовий отвір, а на місці його заднього краю проривається анальний отвір. Третій зародковий листок (мезодерма) виникає із ентодерми.
Загальний план будови тіла первинноротих перебачає двобічну симетрію. Системи органів розміщуються таким чином: на черевній стороні знаходиться центральна нервова система (ортогональна із різними ступенями злиття гангліїв), серце та основна аорта (замкнена, незамкнена кровоносна система, іноді відсутня) знаходяться на спинній стороні, між ними розміщена травна система. Порожнини тіла у різних типів представлені паренхімою, схізоцелем, целомом та міксоцелем. Внутрішній скелет відсутній (у мечохвостів є ендостерніт— справжній внутрішній скелет) Такий план будови із різними ступенями ускладнення чи редукції притаманний для усіх типів первинноротих.
Вториннороті тварини
Єхидна (Tachyglossus aculeatus) представник вторинноротих тварин
Вториннороті об'єднують типи тварин, які характеризуються спільним комплексом ембріологічних ознак. Вони відрізняються від первинноротих тим, що дробіння яйця у вторинноротих радіальне й переважно не детерміноване. У результаті дробіння утворюється бластула, яка шляхом інвагінації перетворюється на двошарову гаструлу. Бластопор гаструли не перетворюється на ротовий отвір, як у первинноротих. На його місці виникає анальний отвір, а рот (вторинний) утворюється на черевній стороні протилежного (переднього) кінця тіла. Третій зародковий листок (мезодерма) виникає шляхом утворенням бічних виростів ентодермального кишківника— ентероцельним способом.
План будови тіла є «дзеркальним» до такого ж у первинноротих. У вторинноротих центральна нервова система (трубчаста із різними ступенями ускладнення) розміщена зі спинної сторони, під нею внутрішній скелет, нижче якого— травна система, а на черевній стороні серце (іноді відсутнє) та основні артерії і вени. Кровоносна система замкнена. Порожнина тіла представлена лише целомом, який закладається у вигляді трьох мішків (залишки радіальної сиеметрії тіла). Симетрія тіла двох типів: радіальна (Голкошкірі) та білатеральна (Хордові, Покривники, Напівхордові).
Надорганізмові живі системи
Надорганізмовими живими системами є угруповання окремих живих організмів або їх груп базованих на різного типу складності взаємодіях, які взаємопов'язані між собою за принципом емерґентності. Розрізняють такі надорганізмові системи[44]:
Моноцен
Моноцен— це найпростіша екосистема, яка ґрунтується на взаємодії будь-якого індивідуальногоорганізму із факторамидовкілля (монотопу) та іншими індивідуальними організмами. У межах моноцену реалізується найпростіший рівень екологічних взаємодій: акція та реакція. Система організм—середовище носить двосторонній характер— середовище впливає на організм; організм впливає на середовище. Кожен із компонентів цієї системи взаємозмінюється. Наприклад, поодиноке дерево у лузі внаслідок виділення речовин через корені, через листковий опад, через затінення ґрунту тощо, спричинює виникнення відмінних від луки екологічних умов у найближчому до нього середовищі, з однієї сторони. З іншої сторони, добра інсоляція сприяє розвитку великої і крислатої крони, що є неможливим у лісі, де проявляється конкурентна боротьба за світло.
Термін моноцен запропонований німецьким зоологом К. Фредеріксом у 1958 році[44].
Розрізняють біотичні й абіотичні фактори, які впливають на індивідуальний організм.
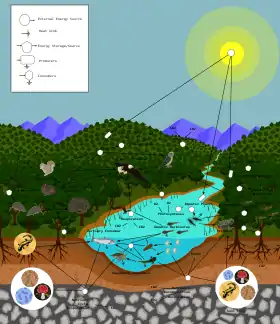
Схема трофічних взаємодій у біоценозіПлеоцен— надорганізмова жива система, яка представлена сукупністю усіх живих організмів (рослини, тварини, гриби, бактерії, археї, віруси та ін.) на певній ділянці суші чи водойми (річки, озера, моря, океани), що співвіснують разом і пов'язані між собою трофічними ланцюгами, потоками матерії та енергії. За своєю суттю плеоцен є сукупністю різновидових демоценів, які постійно взаємодіють між собою (хижацтво, паразитизм, симбіоз та ін.), а також коеволюціонують. Такі біологічні угруповання називають біоценозом, а середовище їх існування— біотопом.
Біоценотичний рівень характеризується переважанням дизкоопераційних коакцій (хижацтво, міжвидова конкуренція тощо) між популяціями різних видів. Тут домінують зв'язки експлуаційного типу, коли одна популяція використовує іншу для свого виживання. За винятком автотрофів, популяції усіх інших видів є одночасно експлуататорами і експлуатованими. Наприклад, рослини експлуатуються пасовищними хижаками (сарни, олені, зайці, миші та ін.), їх експлуатують справжні хижаки (вовки, соколи, леви тощо), а паразити (це також форма хижацтва) експлуатують і пасовищних, і справжніх хижаків. Поруч із експлуатаційними зв'язками важливе місце займають конкуренційні та коопераційні зв'язки.
Біосфера— це глобальна екосистема, яка охоплює усю планету і характеризується глобальними геохімічними циклами. Структурно, біосфера складається із сукупності взаємопов'язаних між собою біомів, кожен з яких об'єднує однотипні біогеоценози, що розподіляються відповідно до кліматичних зон.
Термін біосфера запропонований австрійським геологомЕдуардом Зюсом у 1875 році.
Біосфера, як частина планети населена живими істотами, об'єднує нижній шар атмосфери, гідросферу та верхній шар літосфери. Частина атмосфери населена живими організмами називається аеробіосферою, літосфери— літобіосферою, гідросфери— гідробіосферою[46].
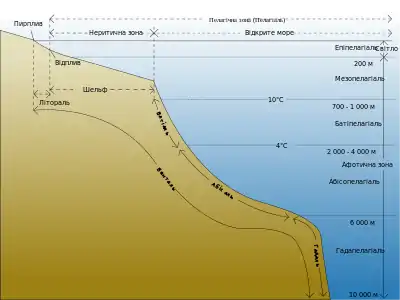
Частина атмосфери, населена живими організмами, називається аеробіосферою[46]. Однак, вона заселена неоднорідно— чим ближче до поверхні Землі, тим більше живих організмів трапляється у атмосфері. Живі організми не можуть постійно перебувати у товщі повітря, оскільки, за своєю суттю, є субстратними істотами, тому вони тут не мешкають, а лише тимчасово перебувають у товщі атмосфери. У атмосфері перебувають птахи, рукокрилі, комахи, гриби, бактерії, віруси, пилокрослин, біогенні речовини тощо.
Приземна частина атмосфери, шаром 50—300м, наповнена живими організмами, є своєрідним екотоном між аеро-, терра- та гідробіосферами. Над ним знаходиться шар атмосфери (300—6000м), куди спорадично можуть підійматися перелітні мігруючі тварини (птахи, комахи тощо)— це тропобіосфера. Над нею знаходиться альтобіосфера (6000—15000м), де максимальна температура ніколи не перевищує 0°C. Сюди повітряними течіями заносить випадкові організми. У верхній частині альтобіосфери знаходиться озоновий шар. Над ним знаходиться парабіосфера, де живі організми відсутні[46].
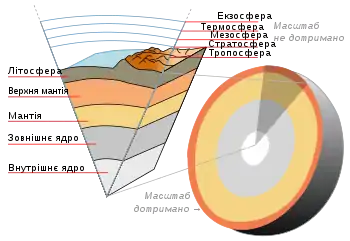
Нижня межа в літосфері
Будова літосфери
Частина літосфери, що населена живими організмами, називається літобіосферою[46]. Її товщина, нижче поверхні ґрунту, становить від 3,5км на дні океану, і до 7,5км на континентах. Вона визначається температурою переходу води в пару і температурою денатураціїбілків, однак в основному поширення живих організмів обмежується вглиб кількома метрами.
Найбільш щільно заселеною живими організмами є поверхня літосфери— террабіосфера,— яка включає рослинний покрив (фітосферу) та кількаметровий ґрунтовий шар (педосферу). У межах террабіосфери сконцентровано більше половини усіх видів живих організмів на Землі. Нижче педосфери, до глибини 1км, лежить гіпотеррабіосфера, яка населена бактеріями та археями, і як виняток, евкаріотами. У глибині літосфери, нижче 1км, відсутній кисень, де проживають хемотрофніанаероби— це теллуробіосфера, завглибшки 1—5км. Нижче неї— до глибини 7—7,5км розміщена гіпобіосфера. Із гіпобіосферою часто пов'язують гіпотезу «Гарячої глибинної біосфери»Томаса Ґолда та біогенне продукування природного газу й нафти археями[47].Гіпобіосфера населена здебільшого термофільними— температура тут сягає 80—105°C— хемотрофними мікроорганізмами. На глибині понад 7,5—8км— метабіосфера,— на сьогодні, життя не виявлено.
Нижня межа в гідросфері
Гідросфера— це єдина географічна оболонка Землі цілковито і повсюдно заселена живими організмами, яка називається гідробіосферою[46]. Гідробіосферу розподіляють на прісноводну частину— аквабіосферу, та океанічну— маринобіосферу. Якщо континентальніпрісні та солоні водойми є порівняно неглибокими— максимальна глибина становить 1642м, яка зареєстрована в озеріБайкал[48], то максимальна глибина океану становить 10 971 ± 10м у Маріанській западині (захід Тихого океану)[49].
Маринобіосферу розподіляють на три великі зони: фото-, дисфото- та афотобіосфери[46]. Фотобіосфера океану щільно населена організмами. Тут відбуваються активні процеси утворення первинної біомаси та фотосинтезу— морські фотосинтетики виділяють понад половину усього річного об'єму кисню на Землі[50]. Світло коротких довжин хвилі проникає до глибини приблизно 200 м— це межа фотобіосфери, нижче неї знаходиться дисфотобіосфера— де фотосинтез у його «класичному» вигляді не відбувається, проте, він можливий у бактерій GSB1, які використовують слабке світло від гідротермальних джерел, а не сонця[51]. Однак, у товщу дисфотобіосфери частково проникає світло довгохвильової частини спектру, що дозволяє фотосинтез і поширення червоних водоростей до глибин 250—300м. Нижня межа дисфотобіосфери пролягає на глибині близько 1км, куди світло не проникає взагалі. Звідси розпочинається афотобіосфера, яка досягає найглибших западин світового океану. Первинна біомаса у афотобіосфері утворюється за рахунок активних процесів хемосинтезу різних типів.
Біотичне розмаїття на Землі
Розмаїття живих організмів
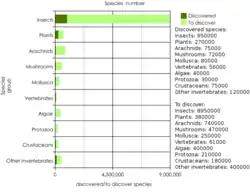
Оцінка співвідношення описаних та невідкритих видів живих істот на Землі (при загальній чисельності близько 10млн видів)
Оцінки вчених щодо ймовірного числа сучасних видів живих істот на Землі розходяться в межах від 10 до 100 мільйонів, що пов'язано із використанням різних розрахункових методик. Загальна кількість відомих видів станом на 2010 рік становила 1,7 мільйона[52], включаючи: зелених рослин— 321212 видів, грибів та лишайників— 48496, бурих водоростей— 3067, тварин— 1250000 (безхребетних— 1305250, з них комах— 1000000; хребетних— 62305, з них риб— 31300 видів).
На еволюцію та розвиток розмаїття біологічних видів існує два кардинально протилежні погляди: нелімітоване і лімітоване видоутворення[53]. Згідно з уявленням про нелімітоване видоутворення, сучасне біорозмаїття сформувалось в результаті комбінації віку певної таксономічної групи та швидкості видоутворення (мінус швидкість вимирання)[54][55]. Уявлення про лімітоване видоутворення передбачає, що виникнення нових видів можливе лише за умови наявності (або вивільнення) доступних ресурсів чи екологічної ніші[56]. Гіпотетично, біологічне розмаїття на Землі повинне зберігатись на приблизно одному і тому ж рівні з часу формування суходільних екосистем (за винятком глобальних вимирань)[57]. Сучасні комп'ютерні моделювання обох гіпотез показують, що еволюція
видового розмаїття має певний ліміт (верхню межу) і відбувається не з однаковою швидкістю[53]
Сучасне розмаїття видів живих істот на Землі не перевищує 1% від числа видів, котрі існували за усю історію нашої планети[58].
Типовий ландшафт тундри (Аляска, США)Тундра— трав'янистийбіом з домінуванням злаків, осок та лишайників, що охоплює північні полярні регіони суходолу вище 60° північної широти. З півдня тундра межує із лісотундрою і тайгою. Розповсюдження тундрових екосистем на материках є нерівномірним— на заході вони завжди зміщені в сторону високих широт (65-68° пн. ш.), а на сході— нижчих (57-59° пн. ш.). Це пояснюється пом'якшенням кліматичних умов західними вітрами, що приносять тепло з океану. Екологічні умови тундри дуже суворі середня температура становить −28°C, кількість опадів 150—250 мм/рік (рівнозначно пустелі), швидкість вітру може досягати 48-97 км/год., період вегетації 60 днів. Тут існує лише дві пори року: зима (полярна ніч) та літо (полярний день)[6].
Розрізняють «високу» та «низьку» тундри. «Висока» тундра— арктична пустеля, що охоплює території із добре вираженою рельєфністю (гори, горби), вона, практично, повністю позбавлена рослинності. «Низька» тундра являє собою рівнинні території (здебільшого узбережні), вкриті товстим шаром нерозкладеної органіки, яка накопичується в результаті сповільненого мікробного розкладу; трав'янистою рослинністю та лишайниками. Для низької тундри притаманні численні водойми і болота[6]. Продуктивність тундри становить 140 г/м² або у планетарному масштабі 1,1×109 тон чистої первинної продукції[23].
Видове багатство тундри є дуже низьким— тут налічується близько 1700 видівсудинних рослин та 48 наземних ссавців, окрім того щорічно, на гніздовий період, в тундру прилітають мільйонні зграї водоплавних птахів[60]. Найхарактернішими тваринами тундри є північні олені (у Євразії), карибу (у Північній Америці), вівцебики, білі ведмеді, полярні зайці, песці, лемінґи, полярні сови, полярні куріпки та ін.
Типовий ландшафт тайги (Аляска, США)Тайга (північні хвойні ліси)— лісовийбіом утворений здебільшого темнохвойними видами дерев смерекою та ялицею, а також сосною та модриною, зі слабо вираженим підліском та чагарничково-трав'янистим ярусом. Тайга охоплює значні території північної півкулі між 45° та 65° пн. ш., вона становить 29% усіх лісів Землі[61]. Як і тундра, тайга простягається з північного заходу на південний схід Євразії та Північної Америки. Південна межа тайги на заході проходить по паралелі 58-60° пн. ш., а на сході— 45-48° пн. ш., що також зумовлюється теплими повітряними масами, які приносяться західними вітрами із океану (Тихого і Атлантичного). Клімат тайги континентальний із жарким і сухим літом та дуже холодною зимою. Середньорічні температури складають від −3°C до +1°C, кількість опадів є дуже низькою— 200—700 мм/рік (зіставно зі степовою зоною). Літо триває 2-3 місяці, а зима— 5-7. Вегетаційний період триває 90-130 днів, а безморозний період— 80-140 днів.
Ґрунти бідні, кислі, із підзолистим профілем, що зумовлено повільним розкладом глиці в умовах холодного клімату.
Для тайгового лісу притаманна висока зімкнутість крон, яка практично не пропускає сонячні промені під шатро, тому чагарники і трави відсутні, а переважають мохоподібні. Незважаючи на слабкий розвиток нижніх ярусів лісу, у тайгових екосистемах суцільний покрив хлорофілу зберігається цілий рік[6], що, незважаючи на низькі температури, забезпечує високий рівень річної продукції: згідно з Ю. Одумом— близько 28-30 кг/м²/рік)[6], згідно з Дж. Равеном— 800 г/м² або у планетарному масштабі 9,6×109 тон чистої первинної продукції[23].
Флора і фауна тайги досить різноманітна, особливо у порівнянні із тундрою, наприклад, видове багатство Східносибірської тайги представлене більш, ніж 2300-ми видами рослин; на території Центрально-Сибірського плато налічується понад 1010 видів судинних рослин[62]. У Північно-Європейській тайзі переважають смерекові ліси; в уральській— світлохвойні соснові ліси; Західному та Східному Сибірах— модринові рідколісся; у канадійській тайзі— смеречняки з модринниками.
У фауні Східносибірської тайги налічується 4 види амфібій, 2 види плазунів, 42 види риб, 203 види птахів та 80 видів ссавців.
Типовий ландшафт листопадного лісу помірного клімату північної півкулі Землі (Чорногорія)Типовий ландшафт листопадного лісу помірного клімату південної півкулі Землі (Нова Зеландія)Листопадні ліси помірного клімату— лісовий біом утворений листопадними листяними видами дерев: дубом, буком, кленом, грабом та іншими, у північній півкулі, а у південній— переважно південнобуком. Листопадні ліси помірного клімату охоплюють території обох півкуль. У північній вони розповсюджені на атлантичному узбережжі Північної Америки, Європі, північному заході Азії (із заходу до 75° сх.д.) та Далекому Сході (вздовж 130° сх.д.). У західних частинах материків вони зміщені до півночі (між 45 і 60° пн.ш.), а у східних— до півня (між 30 і 54° пн.ш.). У південній півкулі ці ліси розповсюджені на півдні Південної Америки (на північ до 50° пд.ш.), південному сході Австралії та Новій Зеландії (південніше 25° пд. ш.). Характерною рисою цього біому є опадання листя на час холодної пори року. Кліматичні умови існування листопадних лісів помірні з добре вираженими чотирма порами року. Середні річні температури становлять +10°C; кількість опадів— 700—1500 мм/рік.
Для листопадового лісу помірного поясу характерна добре виражена і розвинена ярусність: найвищим ярусом є ярус крон видів-едифікаторів, другий ярус представлений кронами видів-співедифікаторів, третій ярус— це підлісок, четвертий— ярус трав і чагарничків. Продуктивність листопадних лісів помірного клімату становить 1200 г/м² або у планетарному масштабі 8,4×109 тон чистої первинної продукції[23].
Найбагатшими із листопадних лісів помірного клімату є далекосхідні третинні реліктові ліси, які збереглись у незмінному вигляді із дольодовикового часу, завдяки їх ізольованості і сталості тутешнього клімату. Основними лісоутворюючими породами є Quercus liaotungensis, Quercus mongolica, Tilia amurensis, Fraxinus manshurica, до них домішуються Ulmus propinqua, Lespedeza bicolor, Corylus heterophylla, Rhamnus dahuricus, Crataegus pinnatifida, Rosa dahurica та ін.[63] Тут поширені
Європейські листопадні ліси мають вторинне походження і сформувалися уже у післяльодовиковий час, мігрувавши із ізольованих локалітетів— рефугіумів у середземномор'ї. Основними лісоутворюючими породами виступають Fagus sylvatica та Quercus robur за участі Carpinus betulus, Tilia cordata[63].
Листопадні ліси помірного клімату півдня Південної Америки мають реліктове походження й збереглись із мезозойської ери, належачи до антарктичної флори[64]. Основні лісоутворюючі породи: Nothofagus betuloides, Nothofagus pumilio та Nothofagus antarctica.
Листопадні помірні ліси Австралії також належать до антарктичної флори і представлені евкаліптовими та південнобуковими лісами. Звичайними у евкаліптових лісах є Eucalyptus microcorys, Eucalyptus pilularis, Eucalyptus grandis, Eucalyptus cloeziana, Lophostemon confertus та ін. У південнобукових лісах домінують Nothofagus moorei та Eucryphia moorei[65].
Листопадні помірні ліси Нової Зеландії також належать до антарктичної флори і передставлені основними лісоутворюючими породами Nothofagus menziesii, Nothofagus fusca, Nothofagus solandri та ін.[66]Типовий ландшафт ковилового степу (Саратовська обл., Росія)Степи (прерії)— це трав'янистийбіом з домінуванням злаків та різнотрав'я. Степи поширені в глибині материків в умовах з недостатньою кількістю опадів для зростання лісів, та надмірною для утворення напівпустель. У східних районах із низькою кількістю опадів тайга безпосередньо переходить у степ. У західних— листяні ліси поступово замінюються на лісостеп, а ті у степ. Степи поширені між 50-35° пн. ш. Клімат степової зони континентальний із середньорічними температурами +10°C, кількістю опадів— 600—650 мм/рік на заході та 200 мм/рік на сході. Степи також поширені і у лісовій зоні в умовах, де екологічні фактори (високий рівень ґрунтових вод, пожежі) перешкоджають зростанню лісів і сприяють експансії злаків[6].
Продуктивність степів становить 600 г/м² або у планетарному масштабі 5,4×109 тон чистої первинної продукції[23]. Слід зауважити, що біомаса підземної частини рослин степу в кілька разів перевищує масу надземних[6]. Це пояснюється тим, що корені більшості багаторічних злаків у клімаксових угрупованнях проникають у ґрунт на глибину до 1,5м.
Степові ґрунти багаті на гумус і представлені чорноземами.
Тваринний світ степів характеризується домінуванням бігаючих (сайгаки, коні, бізони тощо) та риючих (сліпаки, полівки, кроти та ін.) форм ссавців та нелітаючих (дрохви тощо) та наземно гніздних птахів, значною часткою нелітаючих форм комах (дибки, коники, чорнотілки, туруни тощо).
Типовий середземноморський вічнозелений широколистяний субтропічний ліс у ГалілеїВічнозелені широколистяні субтропічні ліси— лісовийбіом утворений вічнозеленими листяними видами дерев: дубом, лавром, фісташкою, фіґою, маслиною, ялівцем, евкаліптом, миртом, суничним деревом, маґнолією, окрою, тамариндом, деякі пальми та ін. Вічнозелені широколистяні субтропічні ліси охоплюють незначні території у обох півкулях: Середземномор'я, ЧилійськийМаторал, Каліфорнію, Капську зону, південно-західну Австралію. Цей тип біому локалізується здебільшого на західних окраїнах материків, повз яких пролягають холодні океанічні течії, що зумовлює специфіку клімату— сухе і спекотне літо та прохолодна (без морозів) і волога зима. На східних окраїнах материків— Японії— вічнозелені широколистяні субтропічні ліси відзначаються вологим і м'яким кліматом.
Вічнозелені широколистяні субтропічні ліси охоплюють близько 10% усього різноманіття рослин на Землі[67].
Продуктивність вічнозелених широколистяних субтропічних лісів становить 1300 г/м² або у планетарному масштабі 6,5×109 тон чистої первинної продукції[23].
Типовий ландшафт савани (Буркіна Фасо)Савани— це трав'янистийрідколіснийбіомтропічного клімату, який характеризується високою кількістю опадів (1000—1500 мм/рік) впродовж дощового періоду та наявністю тривалого посушливого сезону з численними пожежами[6]. Савани охоплюють терени Африки, Австралії, Південної Америки та Мезоамерики. Особливістю ландшафту савани є просторі злакові луки із поодинокими деревами або групами чагарників і термітниками. Савани розташовуються в діапазоні 15°пн.ш.— 15°пд.ш., а в Африці— до 30°пд.ш. Цей біом є перехідною ланкою між пустелями у вищих широтах та дощовими тропічними лісами в районі екватора, у зв'язку з чим опади по території савани розподіляються дуже нерівномірно— їх кількість зростає в напрямку до екватора, і зі сходу на захід.
Продуктивність савани становить 900 г/м² або у планетарному масштабі 13,5×109 тон чистої первинної продукції[23].
Фіторізноманіття саван є дуже низьким— часто на великих територіях домінує один вид злаків та дерев, що зумовлюється посухами та пожежами[6]. У найбільшій на планеті— африканській савані, домінуючими злаками виступають Panicum, Pennisetum, Andropogon, Imperata, а з-поміж дерев переважають акації, баобаби, деревоподібні молочаї та пальми.
Тваринний світ африканської савани є найбагатший на Землі з численними антилопами, зебрами, жирафами, слонами, левами, леопардами, гепардами, гієнами, страусами та іншими представниками сучасної меґафауни.
Типовий ландшафт пустелі (Алтай, Росія)Пустелі
Найбільші пустелі Землі розташовані між 15 і 30 паралелями обох півкуль— Сахара, Аравійська пустеля, Австралійська пустеля та ін. Розповсюдження пустель пов'язане із недостатністю опадів— річна кількість опадів менша 250мм. Це зумовлюється різними факторами, які відносяться до трьох основних категорій: 1. ділянки атмосфери із постійно високим тиском (Північна Африка, Центральна Австралія); 2. локалізація у «дощовій тіні» (пустелі на західних узбережжях материків: Наміб у Африці, Наска у Південній Америці, Каліфорнія у Північній Америці тощо); 3. висотою місцевості (Гобі, Тибет, пустелі Болівії тощо)[6]. Пустелі поділяють на холодні— розповсюджені у помірних або субтропічних регіонах; і гарячі— типові для тропіків.
Пустельна рослинність дуже бідна і представлена трьома біоморфами: однорічниками— ростуть і плодоносять у період випадання дощів (навесні); сукулентами— накопичують воду в товстих стеблах і листках (кактуси, алое тощо); пустельні чагарники— невеликі листопадні колючі кущики із жорстким дрібним листям[6].
Тваринний світ дуже бідний, представлений видами, які можуть довший час обходитись без води, виробляючи із жирових запасів, так звану, «фізіологічну воду», або виробили способи збору води із туману та роси. Здебільшого у пустелях проживають форми тварин, що риють та бігають.
Продуктивність пустель становить 90 г/м² або у планетарному масштабі 1,6×109 тон чистої первинної продукції[23].
Типовий ландшафт тропічного дощового лісу (Квінсленд, Австралія)Тропічні дощові ліси— лісовий біом утворений вічнозеленими листяними видами дерев в умовах надмірного зволоження— 2000—2500мм річних опадів. Особливістю клімату, у якому розповсюджені тропічні дощові ліси, є незначна різниця у коливанні сезонних температур, при суттєвій добовій їх відмінності. Тропічні дощові ліси розповсюджені вздовж екватора в межах 5-10° південної і північної широт. Основні райони їх розповсюдження— басейни Амазонки та Оріноко у Південній Америці, басейн Конґо у Африці та Індокитай, пн.-сх. Австралія та острови Океанії[6].
Дощові тропічні ліси відзначаються дуже добре вираженою ярусністю. Перший ярус (так званий «емерджентний») лісу становлять поодинокі високі дерева, які здіймаються над лісовим шатром на 40-60м; другий— суцільне лісове шатро висотою 25-30м; третій, нижній ярус— розріджений підлісок, який стає густим лише за умови вивалу дерев у першому або другому ярусах. Трав'яний і чагарниковий яруси практично не розвинені і представлені папоротями і пальмами. У тропічному дощовому лісі переважають життєві форми дерев із поверхневою кореневою системою, а також різноманіття ліан.
Екологічною особливістю дощового тропічного лісу є концентрація біогенних елементів у біомасі, а не у ґрунті, як у лісах помірного клімату[68]
Продуктивність тропічних дощових лісів становить 2200 г/м² або у планетарному масштабі 34,7×109 тон чистої первинної продукції[23].
Біотичне розмаїття у дощових тропічних лісах є найвищим на землі, яке становить близько половини усіх відомих на нині видів живих істот. Однією із найбагатших екосистем землі вважаються дощові тропічні ліси у екореґіоні Напо у басейні річки Напо— лівої притоки Амазонки (Еквадор, Колумбія та Перу)[69].
Вертикальна зональність
Пояс циркумбореальних альпійських лук у Альпах, ШвейцаріяПояс центральноафриканських альпійських лук у Гора Кенія, КеніяПояс австралайзійських альпійських лук у Австралійських Альпах, АвстраліяПояс альпійських лук парамо у Андах з деревоподібними Есплетіями, КолумбіяПояс альпійських лук— сукупність структурно простих трав'яних або лишайниковихекосистем з переважанням подушкоподібних низькорослих рослинних біоморф, які розповсюджені над межею лісу у високих горах по всьому світу, тотожні біомутундри. Альпійські луки притаманні також гірським системам тропічних регіонів Землі.
За кліматичними особливостями альпійський пояс відносять до холодної гірської зони. Суми активних температур не досягають 600°С, середньорічна температура близька до 0°С. Загальний період вегетації становить приблизно 90 днів.
Ґрунти оліготрофні, гірсько-лучні, схильні до оторфовіння, через те, що акумуляція органічних решток переважає над їх розкладанням.
•Циркумбореальні альпійські луки розповсюджені у горах Європи, Північної Азії та Північної Амерки на висотах 1800-3000м над рівнем моря. Для екосистем циркумбореальних альпійських лук характерні угруповання ситників (Juncus L.), осок (Carex L.) та костриць (Festuca L.) – це переважно щільно дернинні дрібнозлакові або різнотравні відкриті скельні луки. Поширені також чагарничково-мохово-лишайникові пустища. Різноманітність судинних рослин незначна, більше представлені лишайники й мохи.
•Східноафриканські альпійські луки розповсюджені у Горах Східної Африки: Рувензорі, Гора Кенія, Гора Кіліманджаро та ін. на висотах 4000-4500м над рівнем моря. Східноафриканські альпійські лучні екосистеми утворені домінантними видами з родів безсмертник (Helichrysum) та приворотень (Alchemilla)[70] за значної участі осоки одноколоскової (Carex monostachya A.Rich.), мітлиці вузьколистої (Agrostis trachyphylla Pilg.) та ін.[71] з поодинокими деревоподібними будяками (Carduus L.), жовтозіллями (Senecio L.)дерево-жовтозіллями (Dendrosenecio (Hauman ex Hedberg) B. Nord.) та лобеліями (Lobelia L.).
•Гімалайські альпійські луки поширені у Гімалаях та на Алтаї на висотах 3000-4500м над рівнем моря.
•Австралайзійські альпійські луки розповсюджені у горах Папуа-Нової Ґвінеї, Австралійських Альпах та новозеландськихПівденних Альпах. На Папуа-Новій Ґвінеї альпійські луки притаманні здебільшого для Новоґвінейського Центрального Хребта на висотах понад 3500м над рівнем моря й представлені угрупованнями щучника (Deschampsia Beauvois) і тонконога (Poa L.). В Австралії альпійські луки займають висоти вище 1800м над рівнем моря. Рослинність представлена угрупованнями тонконога (Poa L.) та ін. асоційованих з ним видів трав'янистих рослин. У Новій Зеландії альпійські луки займають висоти близько 1800м над рівнем моря й утворені хіонохлоєю (Chionochloa), хіоногебією (Chionohebe) й аціфілою (Aciphylla).
•Південноамериканські альпійські луки (парамо) розповсюджені у Андах на висотах 3500-4100м над рівнем моря і включає три зони: верхнє парамо (суперпарамо), лучне парамо та нижнє парамо (субпарамо)[72]. Екосистеми утворені угрупованнямикуничника (CalamagrostisAdans.) та костриці (Festuca L.), з поодинокими, заввишки до 2-5м, вічнозеленими деревоподібними айстровими з роду Есплетія (Espeletia Mutis ex Humb. & Bonpl.)[73]. У нижньому парамо чагарники займають домінуюче становище і утворені представниками падуба (Ilex L.), аґератини (Ageratina Spach) та бакхарисами (Baccharis L.).
Субальпійське криволісся із сосни гірської на г. Мала Попадя в Ґорґанах, УкраїнаСхідноафриканське субальпійське гірське криволісся, Гори Рувензорі, УгандаПівденноазійське субальпійське гірське криволісся, г. Кінабалу, Борнео, ІндонезіяПояс субальпійського криволісся— сукупність гірських чагарникових екосистем, утворених корінною чагарниковою сланкою рослинністю, вторинних субальпійських лук (полонини) та чорнично-мохові пустирі, розміщених між альпійським і лісовим гірськими поясами.
Згідно з кліматичним зонуванням пояс приурочений до холодної зони і характеризується кліматичними умовами, подібними до альпійського поясу.
•Циркумбореальні субальпійські гірські криволісся поширені у Альпах, Карпатах, Кавказі, Скелястих Горах та ін. північних гірських системах субальпійський пояс розміщується на висоті від 1300—1500м над р.м. до 2500м над р.м. Тут поширені сосна гірська, вільха зелена, ялівець сибірський, рододендрони, а у горах середземноморського регіону та Північної Африки— деревовидними вересами-еріками ([верес-еріка іспанська
(Erica lusitanica Rudolphi), верес-еріка віникова (Erica scoparia L.) та верес-еріка деревовидна (Erica arborea L.) та ін чагарники високотрав'я та різнотрав'я. Зарості сосни гірської приурочені до торф'янистих і торфовихґрунтів, на яких поширені також чорниця і мохи. Зеленовільшняки приурочені до затінених схилів або улоговин з вологими чи сирими ґрунтами. Тут трав'янистий покрив багатший. Зустрічаються тирличі, безщитники, жовтозілля тощо. Сухіші та освітлені південні схили займають зарості ялівцю сибірського. На місці вирубування субальпійських криволісь утворюються вторинні луки, які у Карпатах називаються «полонини»
•Східноафриканські субальпійські гірські криволісся поширені здебільшого у гірському масиві Рувензорі, а спорадично також у Горах Вірунґа, Горах Мітумба, Абердарських Горах та вершинах Гора Кенія, Гора Кілімаднжаро, на висотах 3000-4000 м над рівнем моря. Криволісся представлені Вересом-Ерікою деревовидною, деревовидними лобеліями (лобелія Декена (Lobelia deckenii), лобелія абердарська (Lobelia aberdarica) та ін.) та дерево-жовтозіллями (дерево-жовтозілля прильодовикове (Dendrosenecio adnivalis), дерево-жовтозілля кенійське (Dendrosenecio keniensis), дерево-жовтозілля кіліманджарське (Dendrosenecio kilimanjari) та ін.).
•Гімалайські субальпійські гірські криволісся поширені у Гімалайських та прилеглих гірських масивах Алтаю та ін., на висотах від 3000м до 4000м над рівнем моря. Рослинність представлена криволіссями із деревовидних рододендронів (наприклад, рододендрод кущовий (Rhododendron arboreumboreum) та ялівців.
•Південноазійські субальпійські гірські криволісся поширені у гірських системах Південно-Східної Азії на висотах 2500-3000м над рівнем моря. На вершині г. Кінабалу, о. Борнео, криволісся представлене заростями дакридіума Ґібса (Dacrydium gibbsiae).
•Австралайзійські субальпійські гірські криволісся поширені у горах Нової Ґвінеї, Австралії, Нової Зеландії. Новоґвінейські криволісся охоплюють проміжок висот між 3000-4000 метрів, представлені чагарниками з родіврододендрон (Rhododendron), чорниця (Vaccinium), копросма (Coprosma), рапанея (Rapanea), зауравія (Saurauia) та ін.
•Південноамериканські субальпійські гірські криволісся поширені у Андах на висотах 3000-3500м над рівнем моря і називаються субпарамо— чагарниковою зоною, що розмежовує альпійські луки— парамо і верхній лісовий пояс. Рослинність складена угрупованнями падуба (Ilex L.), змієкореня (Ageratina Spach.) та бахарису (Baccharis L.).
Пояс тропічних гірських бамбукових лісів
Гірський смеречняк у північній півкулі Землі, Європа
Хвойний пояс із араукарії араукани у Південній півкулі Землі, Арґентина
Пояс гірських хвойних лісів— лісові екосистеми, утворені хвойними деревнимивидами, поширені смугою на висоті 700—1500м над р.м. і вище. У північній півкулі хвойний пояс складають здебільшого представники роду смерека, ялиця та сосна; у південній— араукарія.
Кліматично пояс хвойних лісів приурочений до прохолодної та помірно холодної кліматичних зон із сумою активних температур від 1000 до 1600°С та сумою опадів до 1500 мм/рік. Ґрунти кислі, часто щебнисті.
•Австралайзійські гірські хвойні ліси
Поширені у горах Нової Ґвінеї, Австралії, Нової Зеландії на висотах 2000-3000 метрів над морем. Представлені здебільшого подокарпусами (Podocarpus), дакрикарпусами (Dacrycarpus), дакридіумами (Dacrydium), папуакедрами (Papuacedrus), араукаріями (Araucaria), and лібокедрами (Libocedrus) та ін.
Пояс гірських листяних лісів
•Циркумбореальні гірські листяні ліси
Поширені у горах північної півкулі на висотах 700—1500 м (залежно від широти), де представлені буком (Fagus).
•Австралайзійсько-Південноамериканські гірські листяні ліси
Поширені у горах південної півкулі в межах антарктичної флори на висотах 900—2000 метрів над морем, де представлені лісами з південнобука (Nothofagus).
Екосистеми холодних глибоководних метанових джерел
Екосистеми гарячих суходільних джерел
Екосистеми сольових роп
Екосистеми асфальтових озер
Екосистеми кислотних озер
Виникнення життя на Землі
Теорія самозародження Опаріна-Галдана
Гіпотеза світу РНК
Гіпотеза світу РНК передбачає, що вихідною формою живих істот на Землі були молекулиРНК, які в результаті біохімічноїеволюції розвинулись у ДНК та білки, давши початок білковому життю сучасного типу[4].
Гіпотеза світу РНК заснована на здатності РНК запам'ятовувати, передавати та дублювати генетичну інформацію так само, як це робить ДНК. Окрім того, РНК має каталітичні властивості та може поводити себе як фермент— рибозим (РНК-ензим).
Гіпотеза світу РНК пояснює, як виник складний процес передачі інформації з ДНК на білок, який представлений трьома основними стадіями: транскрипцією— передачею інформації з ДНК на РНК; дозріванням РНК— видаленням зайвих послідовностей нуклеотидів, які не несуть інформації для синтезу білка; трансляцією— передачею інформації з дозрілої РНК на білок. У цій схемі існує один зворотний зв'язок, коли інформація передається з РНК на ДНК, але не відомо жодного випадку передачі інформації з білка на РНК чи ДНК[74].
Панспермія— космогонічна гіпотеза про появу життя на Землі в результаті перенесення з інших планет якихось «зародків життя».
Гіпотеза розумного задуму
Гіпотеза розумного задуму передбачає те, що усі форми життя на Землі та людина були створені вищою, надприродною силою— богом чи богами.
Розвиток життя на Землі
Поява життя на Землі
Перші живі організми сучасного типу з'явились на Землі понад 3,5млрд років тому. Це були одноклітинні істоти з простою організацією клітини. Вважається, що вони були схожими на сучасних прокаріотів: у них було відсутнє ядро та мембранніорганели. Предками перших організмів сучасного типу були, так звані, протобіонти— гіпотетичні істоти, які мали усі риси живих систем, однак, знаходились на нижчій сходинці організації[75]. Найдревніші беззаперечні скам'янілі останки одноклітинних істот датуються часом 3,5млрд років тому[76], а геохімічні сліди живих істот знайдені у відкладах віком 3,8млрд років тому[77],
а також у відкладах віком 3,95млрд років тому[78]
У 2010 році вперше створено організм— паразитичнубактеріюМікоплазму грибоподібну JCVI-syn1.0 (Mycoplasma mycoides JCVI-syn1.0) із цілком синтетичним геномом[79]. Кільцева ДНК-плазміда розміром 1,08 мега-пар нуклеотидів, була спроектована відповідно до оцифрованого геному Мікоплазми грибоподібної і синтезована хімічним шляхом. Синтетичну «хромосому» імплантовано у клітину іншого виду паразитичних бактерій— Мікоплазми козячої (Mycoplasma capricolum), в результаті отримано Мікоплазму грибоподібну JCVI-syn1.0— повноцінно життєздатну бактерію, контрольовану синтетичним геномом.
Перші спроби створити геномсинтетичний евкаріотичний організм здійснені у 2011 році. Здійснено успішні експерименти з імплантації правого плеча 9-ї та лівого плеча 6-ї хромосом у живу клітину Дріжджів пивних .[80]
Генетично модифіковані організми
Генетично модифікованими організми— називають істот, генетичний матеріал яких був змінений шляхом, що не відбувається в природних умовах, на відміну від схрещування або природної рекомбінації. Відповідне формулювання затверджене в статті 2 Директиви 2001/18/ЄС Європейського парламенту та Ради від 12 березня 2001 року про вивільнення у довкілля генетично модифікованих організмів.[81].
Вимирання
Вимирання— це процес поступового або раптового зникнення групи таксонів чи видів, що призводить до скорочення біорізноманіття. Момент вимирання визначається зі смертю останнього представника виду. Понад 99% живих істот, що мешкали на Землі впродовж усієї її історії, вимерли.
Упродовж геологічної історії Землі відбулося 5 глобальних вимирань[82]:
Девонське вимирання— відбулось у пізньому девонському періоді палеозойської ери, приблизно 360млн років тому. Внаслідок цього вимирання зникло близько 30% усіх відомих родин девонської фауни;
Пермське вимирання— відбулось у пізньому пермському періоді палеозойської ери, приблизно 250млн років тому. Наслідком стало зникнення понад 50% усіх родин пермської фауни та флори[83];
Крейдове вимирання— відбулось у пізньому крейдовому періоді мезозойської ери, 65 мільйонів років тому. Внаслідок цього вимирання зникли 17% родин фауни і флори.
У наш час немає наукових доказів існування життя на Марсі. Хоча припускають, що воно там може бути. Ще до початку польотів на Марс він був першим кандидатом на виявлення там позаземного життя.
Непрямі докази існування життя на Марсі
Скам'янілі бактеріоподібні структури були знайдені у марсіанському метеоритіALH84001[84]. Їхні розміри коливаються в межах 20—100 нанометрів. Метеорит ALH84001 знайдено у грудні1984 року в горах Алана, які є східним закінченням Трансантарктичного хребта, на Землі Королеви Вікторії, що в Антарктиді. Однак повторні дослідження структур метеориту ALH84001 показують, що вони мають мінеральне походження, а не органічне[85].
15 січня 2009 року в онлайн-версії журналу «Наука» (Science) опубліковано статтю про виділення метану в атмосферу Марсу[86]. Встановлено, що частка газу метану у складі марсіанської атмосфери становить приблизно 10 мільярдних відсотка. У період марсіанських весни-літа, коли тане мерзлота, його кількість в атмосфері різко зростає, а з похолоданням— зменшується. Джерелом метану можуть виступати мікроорганізми— на Землі 90% усього метану утворюється метаногенними мікроорганізмами.
Європа— це найменший галілеєвий супутник Юпітера, який дуже схожий на планету-океан, яка вкрита шаром води товщиною близько 90—100км і замерзлою поверхнею. В океані спостерігаються припливи до 30м, тепла від яких достатньо для підтримки частини океану в рідкому стані. Припускають, що підлідний океан має термальні джерела і поблизу них може існувати життя. Інші вчені вважають, що океан Європи «мертвий», оскільки містить багато отруйних речовин.
Thomas S. Duval Self-Awareness and Causal Attribution, p. 1, Springer, 2001
Miller, Jason (2009). «Minding the Animals: Ethology and the Obsolescence of Left Humanism». American Chronicle.
Povinelli, Daniel; de Veer, Monique; Gallup Jr., Gordon; Theall, Laura; van den Bos, Ruud (2003). «An 8-year longitudinal study of mirror self-recognition in chimpanzees (Pan troglodytes)». Neuropsychologia 41 (2): 229—334.
Joshua M. Plotnik, Frans B.M. de Waal, and Diana Reiss (2006) Self-recognition in an Asian elephant. Proceedings of the National Academy of Sciences 103(45):17053-17057
Prior, Helmut; Schwarz, Ariane; Güntürkün, Onur; De Waal, Frans (2008). «Mirror-Induced Behavior in the Magpie (Pica pica): Evidence of Self-Recognition». PLoS Biology (Public Library of Science) 6 (8)
Stanley Coren. How dogs think.
Archer, John (1992). Ethology and Human Development. Rowman & Littlefield.
Lovelock, James (2007). The Revenge of Gaia: Why the Earth Is Fighting Back— and How We Can Still Save Humanity. Santa Barbara CA: Allen Lane;
Lovelock, James (2001). Homage to Gaia: The Life of an Independent Scientist. Oxford: Oxford University Press;
Царик Й. В., Царик І.Й.Консорція як один із базових рівнів біологічного різноманіття // Карпатський регіон і проблеми сталого розвитку: Матеріали конф. – Рахів, 1998. – С. 303—304.
Царик Й. В., Царик І.Й.Консорція як загальнобіотичне явище // Вісник Львів. ун-ту. Серія біологічна. – 2002. – вип. 28. – С. 163—169.
Raven, Johnson Biology.— MGH, 2002.— 6 ed.— 1239 p.
Richardson and Keuck, "Haeckel's ABC of evolution and development, " p. 516
Sinkovics, J; Harvath J, Horak A. (1998). «The Origin and evolution of viruses (a review)». Acta microbiologica et immunologica Hungarica (St. Joseph's Hospital, Department of Medicine, University of South Florida College of Medicine, Tampa, FL, USA.: Akademiai Kiado) 45(3-4): 349-90;
Lindquist S, Krobitsch S, Li L, Sondheimer N (February 2001). «Investigating protein conformation-based inheritance and disease in yeast». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 356 (1406): 169-76
Brown DR, Qin K, Herms JW, et al. (1997). «The cellular prion protein binds copper in vivo». Nature 390 (6661): 684-7.
Simonite T. Protists push animals aside in rule revamp// Nature.— 438.— (November 2005) (7064): 8-9.
Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П.О.Ботаніка. Водорості та гриби.— Київ, 2004.
Щербак Г. Й., Царичкова Д. Б., Вервес Ю.Г.Зоологія безхребетних. Том 3.— К.: Либідь, 1997
Кучерявий В.П.Екологія.— Львів, 2001.— 500°C.
Fritz Schwerdtfeger Demökologie. Struktur und Dynamik tierischer Populationen, Band 2, 1968
РеймерсН.Ф. Экология (теории, законы, правила, принципы и гипотезы). М.: Журнал «Россия Молодая», 1994.— 367 с.
Gold T. The Deep, Hot Biosphere 1992 Proceedings of the National Academy of Sciences volume 89 issue 13: 6045-6049
A new bathymetric map of Lake Baikal. INTAS Project 99-1669. Ghent University, Ghent, Belgium; Consolidated Research Group on Marine Geosciences (CRG-MG), University of Barcelona, Spain; Limnological Institute of the Siberian Branch of the Russian Academy of Sciences, Irkutsk, Russian Federation; State Science Research Navigation-Hydrographic Institute of the Ministry of Defense, St. Petersburg, Russian Federation. Morphometric data.
McPeek M. A, Brown J. M (2007) Clade age and not diversification rate explains species richness among animal taxa. Am Nat 169: 97–106.
Ricklefs R (2007) Estimating diversification rates from phylogenetic information. Trends Ecol Evol 22: 601—610.
Van Bocxlaer I, Loader S. P, Roelants K, Biju S. D, Menegon M, et al. (2010) Gradual adaptation toward a range-expansion phenotype initiated the global radiation of toads. Science 327: 679—682.
Cody, M.L. (1986). «Diversity, rarity, and conservation in Mediterranean-climate regions». In Soulé, M.E. Conservation biology: the science of scarcity and diversity. Sunderland, Massachusetts, USA.: Sinauer. pp. 122—152.
Yeoman, G. (1989) Africa's Mountains of the Moon: Journeys to the Snowy Sources of the Nile. Elm Tree Books, London.
Mizuno, Kazuharu (1998). «Succession Processes of Alpine Vegetation in Response to Glacial Fluctuations of Tyndall Glacier, Mt. Kenya, Kenya». Arctic and Alpine Research (INSTAAR, University of Colorado) 30 (4): 340—348.
Luteyn, James L. (1999). Páramos: A Checklist of Plant Diversity, Geographical Distribution, and Botanical Literature. Bronx, New York: New York Botanical Garden Press.
Zimmer C (August 2009). «Origins. On the origin of eukaryotes». Science 325 (5941): 666-8.
Altermann, W. and Kazmierczak, J. (2003). «Archean microfossils: a reappraisal of early life on Earth». Res Microbiol 154 (9): 611-7.
Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison, T.M., Nutman, A.P. and Friend, C.R.L. (November 1996). «Evidence for life on Earth before 3,800 million years ago». Nature 384 (6604): 55-59.

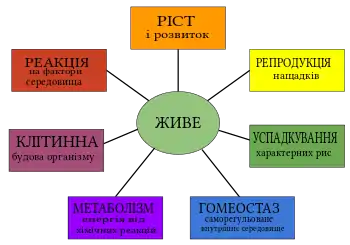




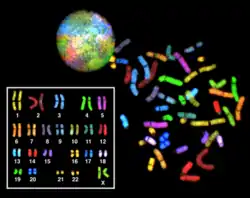
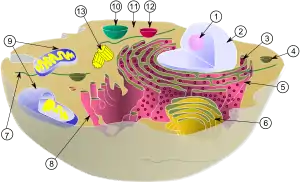



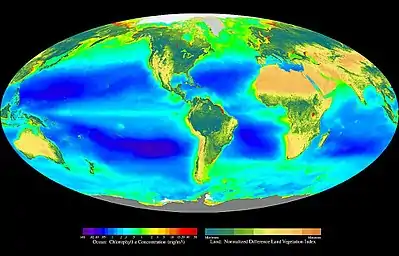
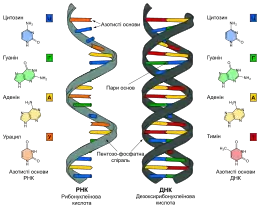
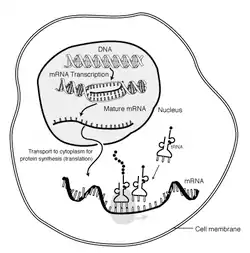
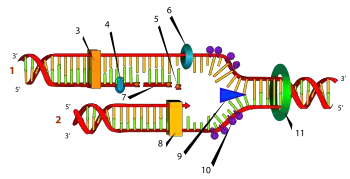

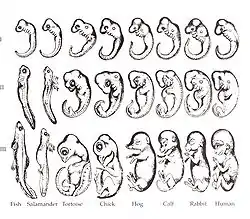
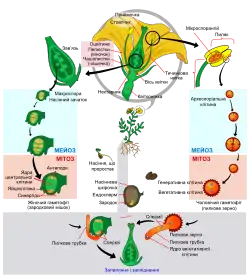
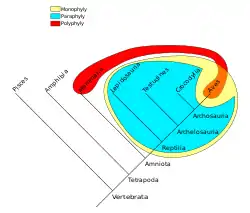
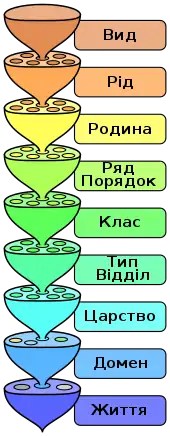
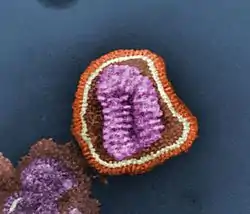

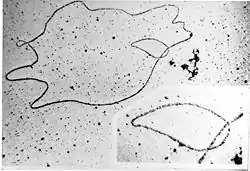
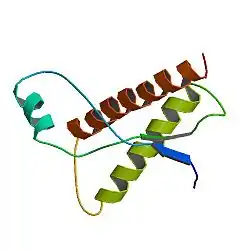
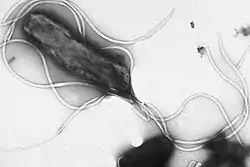









.jpg.webp)
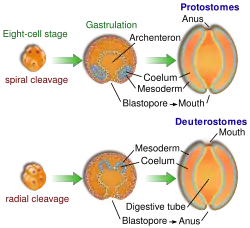
.jpg.webp)