Базидія
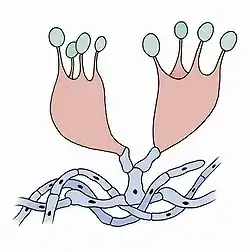
Бази́дії (лат. basidia, однина basidium) — коротко-булавоподібні клітини гіменію, на верхівці яких утворюються чотири стеригми, кожна з однією спорою. Зрідка на базидії утворюються дві спори.
| Базидії |
|---|
 Базидії |
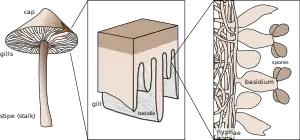
Будова
Основними компонентами базидії є:
тіло базидії — мішкоподібна або циліндрична структура, одноклітинна або розділена на клітини — гаплоцити;[1]
стеригми — конусоподібні вирости, що утворюються на верхній або бічній поверхні тіла базидіїї;[2]
базидіоспори — мейоспори, що утворюються на кінцях стеригм та відділяються від них (іноді — разом із ними) після дозрівання[3]
Статеве спороношення базидіоміцетів відбувається через утворення екзогенних спор на особливих виростах — базидіях. Оскільки статевий процес здійснюється шляхом злиття двох вегетативних клітин гаплоїдного міцелію, що виростає з базидіоспор і складається з одноядерних клітин, для утворення базидій потрібно злиття чоловічого та жіночого гіфів. Об'єднаний в результаті дикаріонний міцелій може утворити плодові тіла з базидіями, хоча у деяких видів виникають прямо на міцелії. На маленьких шипоподібних виростах стеригмах утворюється 2-4 базидіоспори. Розсіювання спор відбувається шляхом активного відкидання, через підняття в базидії клітинного тиску у наслідок гідролізу глікогену. Відкинута спора підхоплюється потоками вітру. У закритих плодових тілах (гастероміцетів) спори не так активно відкидаються, а розсіювання відбувається через порушення зовнішньої оболонки.
Базидії різняться будовою і можуть бути булавоподібні, одноклітковою - холобазидія, складеною з двох частин — нижньої та верхньої — гетеробазидія, складеною з 4 клітин по боках яких утворюються спори — фрагмобазидієя. Через будову базидій безидіоміцети поділяються на 3 класи: холобазидіоміцети, гіменоміцети, гастероміцети.[4]
Класифікація Базидій
Тіло базидії — це складна структура зі своєрідним онтогенезом. У більшості грибів воно утворюється з термінальної клітини генеративної гіфи й лише зрідка — з її бічного відростка. Згідно з розташуванням на генеративній гіфі, базидії поділяють на три типи:
— апікальні базидії — паралельні до осі генеративної гіфи й утворюються з її термінальної клітини;
— плевробазидії — перпендикулярні до осі генеративної гіфи й утворюються з його бічного відростка; при цьому генеративна гіфа продовжує рости й може формувати нові базидії;
— подобазидії — перпендикулярні до осі генеративної гіфи й утворюються з її бічного відростка; на відміну від плевробазидій, генеративна гіфа припиняє свій ріст, отже базидія, що утворюється, відрізняється від апікальної лише своєю викривленою основою.
Онтогенетичною попередницею базидії є базидіоль — клітина дикаріотичного міцелію, в якій відбувається каріогамія й утворюється диплоїдне ядро. Невдовзі після утворення диплоїдне ядро підлягає мейозу, який може відбуватися як у самій базидіолі, так і в новій структурі, що проліферує із неї. Залежно від цього виокремлюють два типи базидій:
— гомобазидії — утворюються безпосередньо з базидіолі після проходження в ній мейозу;
— гетеробазидії — утворюються de novo шляхом проліферації базидіолі. Диплоїдне ядро мігрує у виріст, що утворюється, і вже в ньому підлягає мейозу.
Як уже було сказано, тіло базидії (незалежно від того, чи йде мова про гомобазидію, чи про мета-частину гетеробазидії) може складатися з однієї клітини або бути розділеним на кілька (зазвичай чотири) окремі клітини. Із цього погляду виокремлюють:
— голобазидії — не мають септ; їхнє тіло являє собою одну клітину;
— фрагмобазидії — розділені септами на чотири клітини-гаплоцити.
Останнім часом набула поширення точка зору, що фрагмобазидії є більш давнім типом спороношення, ніж голобазидії. Септи виникають унаслідок мейозу диплоїдного ядра (цитокінез у більшості еукаріотів відбувається відразу ж після каріокінезу), проте не мають у базидії серйозного функціонального навантаження. Як наслідок, у ході еволюції відбувається їхня поступова редукція. Відомий ряд перехідних базидій, що мають часткову перегородку.
Голобазидії не слід плутати з гомобазидіями. Схожість цих назв часто призводить до непорозумінь, тим більше, що «класичні» базидії, характерні, наприклад, для шапинкових грибів, є одночасно і гомобазидіями (утворюються безпосередньо з базидіолі), і голобазидіями (не розділені септами).
Розташування септ у тілі базидії залежить від того, в якій площині відбувається мейоз (точніше — друге ділення мейозу) диплоїдного ядра базидіолі. Якщо площина ділення паралельна до поздовжньої осі базидіолі, то утворювані ядра також розміщуватимуться паралельно до неї, а септи, що їх розділяють,— навпаки, перпендикулярно. Можливий і протилежний випадок. Виходячи із цього, виокремлюють два типи базидій:
— хіастобазидії (хіастичні базидії) — площина мейозу перпендикулярна до поздовжньої осі базидіолі, а септи паралельні до неї;
— стихобазидії (стихічні базидії) — площина мейозу паралельна до поздовжньої осі базидіолі, а септи перпендикулярні до неї (у фрагмобазидіях) або відсутні (у голобазидіях).
Слід зазначити, що на тілі хіастобазидій іноді можуть повторно утворюватися поперечні септи (Exobasidium, Tilletia), проте при цьому в спороутворенні бере участь тільки верхівкова клітина, яка залишається неподіленою, тому такі базидії відносять до холо-типу.
Сучасні автори (наприклад, Swann et al., 2001) указують на те, що стихобазидії є архаїчним типом спороношення. На ранніх стадіях розвитку вони нагадують аски і, ймовірно, утворилися від подібних до них структур, властивих спільним предкам Ascomycota та Basidiomycota. Проте екзогенне спороутворення призвело до того, що окремі спори в межах стихобазидій опинилися в нерівних умовах: верхнім із них простіше залишити плодове тіло, аніж нижнім. Для компенсації цього ефекту стихобазидії іноді набувають Г-подібної форми (Eocronartium), проте лише перехід до хіастобазидій забезпечив остаточне розв’язання проблеми.
Розглянуті підходи до класифікації базидій можу бути об’єднані в одну систему, згідно із якою існує сім основних типів базидій:
— Agaricus-тип — хіастичні гомо-голобазидії;
— Tulasnella-тип — хіастичні гетеро-голобазидії;
— Tilletia-тип — хіастичні гетеро-голобазидії з теліоспорами;
— Tremella-тип — хіастичні гетеро-фрагмобазидії;
— Auricularia-тип — стихічні гомо-фрагмобазидії;
— Septobasidium-тип — стихічні гетеро-фрагмобазидії;
— Ustilago-тип — стихічні гетеро-фрагмобазидії з теліоспорами.
Кожен з перерахованих типів базидій має безліч варіантів, що відрізняються низкою додаткових ознак: кількістю базидіоспор, способом їхнього відділення від базидії, положенням базидії на генеративній гіфі, формою стеригм і тіла базидій, розташуванням септ на базидіях, базидіоспорах і теліоспорах тощо.[5]
Галерея

.jpg.webp) Clavulina coralloides
Clavulina coralloides Psilocybe aztecorum
Psilocybe aztecorum
Примітки
- Akulov, O. V.; Акулов, О. В. (2007). Zahalʹna mikolohii︠a︡. Kharkiv: Osnova. ISBN 978-966-495-040-1. OCLC 664248269.
- Akulov, O. V.; Акулов, О. В. (2007). Zahalʹna mikolohii︠a︡. Kharkiv: Osnova. ISBN 978-966-495-040-1. OCLC 664248269.
- Akulov, O. V.; Акулов, О. В. (2007). Zahalʹna mikolohii︠a︡. Kharkiv: Osnova. ISBN 978-966-495-040-1. OCLC 664248269.
- Горленко, М.В. (1976). Жихнь растений в 6 томах. Москва: Просвещение. с. 479. - С. 227
- Akulov, O. V.; Акулов, О. В. (2007). Zahalʹna mikolohii︠a︡. Kharkiv: Osnova. ISBN 978-966-495-040-1. OCLC 664248269.
Зображення
- http://bio.1september.ru/2000/40/5.gif
- https://web.archive.org/web/20061109000951/http://www.cultinfo.ru/fulltext/1/001/009/001/228465915.jpg
{kind=link}
{kind=link}