Гіперцикл
Гіперцикл — система, яка складається з кількох автокаталітичних одиниць, що пов'язані між собою циклічним зв'язком. Гіпотетичний етап біохімічної еволюції живих організмів. Автор теорії гіперциклів М. Ейген розглядав гіперцикл як білковонуклеотидний комплекс, в якому білок каталізує реплікацію РНК, а РНК в свою чергу є матрицями для синтезу білка.
Короткий виклад
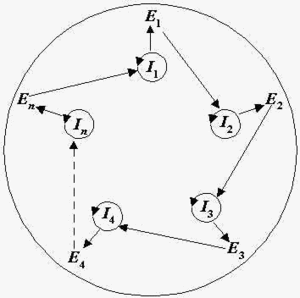
В кінці 1970-х років М. Ейген і П. Шустер запропонували модель гіперциклів. В гіперциклі до ланцюжків РНК додаються ланцюжки амінокислот — білок, які виконують певні каталітичні функції і разом з ланцюжками РНК формують цілісну систему молекул, що кооперативно взаємодіють. Образно кажучи, в гіперциклі ланцюжки РНК кооперуються, але не самі, а з допомогою примітивних поліпептидних ферментів (див. рисунок).
В гіперциклі РНК і ферменти кооперуються наступним чином. Існують РНК-матриці (Ii); і-та РНК кодую і-й фермент Еі (i=1,2,…,n); ферменти циклічно каталізують реплікацію РНК, а саме фермент E1 каталізує реплікацію І2, Е2 каталізує реплікацію І3, …, En каталізує реплікацію І1. Крім того, згадані макромолекули кооперативно забезпечують трансляцію, так що інформація, яка закодована в РНК транслюється в структуру ферментів, аналогічно звичайному механізму трансляції в живих клітинах. Циклічна організація гіперциклу забезпечує його структурну стабільність. Загалом, ферменти могли допомагати підвищенню точності копіювання, в результаті збільшувалась кількість інформації, яку могли передавати потомкам такі структури. Таким чином модель гіперциклів інтерпретує гіпотетичну стадію еволюції, яка могла відбуватись після стадії квазівидів. Схема роботи гіперциклу ще дуже далека від молекулярно-генетичної схеми самовідтворення в живій клітині, однак гіперцикл — це певний крок до живої клітини, порівняно з квазівидами, який вже включає кооперацію між полінуклеотидами і білками.
Об'єктом відбору у еволюції є квазівид — розподіл (генотипічно) споріднених реплікативних одиниць, згрупованих навколо копії (або виродженої множини копій), які відповідають фенотипу із максимальною селективною цінністю. Інформаційний зміст цієї домінуючої копії — виражене у числі символів (нуклеотидів) на одну реплікативну одиницю — обмежений:


де — перевершення домінуючої копії, тобто середнє селективне перевершення над видами іншої частини розподілу, — середня якість копіювання символу. Перевищення цього порогу інформаційного змісту викликає катастрофу помилок, тобто розпад інформації через постійне накопичення помилок. Для досягнення стійкого інформаційного порядку декількох тисяч нуклеотидів потрібно віднайти дуже складний апарат ферментативної реплікації. Такої кількості нуклеотидів як раз достатньо, щоб закодувати декілька білкових молекул, як у нині існуючих РНК-містких фагів. Фізичні властивості, внутрішньо притаманні нуклеїновим кислотам, припускають відтворюване накопичення інформації у кількості, яке не перебільшує 50-100 нуклетидів. Однак наявність певних чинників середовища, наприклад каталітичних поверхонь або навіть белковоподібних попередників ферментів, може значно змістити ці оцінки. Фізичні властивості, притаманні нуклеотидам, дають можливіть дискримінувати комплементарні й некомплементарні нуклеотиди. Існує різниця у вільних енергіях для дискримінації між комплементарними й некомплементарними парами основ.


Дарвінівській еволюції передував багатоетапний процес молекулярної еволюції, який привів до створення унікального апарату клітини, яка використовує універсальний код. Цей код встановився не тому, що він був єдино можливим, а тому, що тут працював своєрідний механізм відбору «раз і назавжди», причому цей процес міг початися з будь-яких кодових відповідностей. Відбір «раз і назавжди» є наслідком циклічної організації. Аналіз механізмів відтворення макромолекул дозволяє думати, що наявність каталітичних гіперциклів — це мінімальна вимога для виникнення макромолекулярної організації, здатної накопичувати, зберігати й обробляти генетичну інформацію.
Каталітичний гіперцикл — це система, яка пов'язує автокаталітичні, або самовідтворювані одиниці одне з одним шляхом циклічного зв'язку. Інтермедіати, як самовідтворювані одиниці самі є каталітичними циклами. Процес реплікації прямо або опосередковано стимулюється додатковими специфічними зв'язками між різними реплікативними одиницями. Реально такі зв'язки можуть забезпечуватися білками — продуктами трансляції передуючих циклів відтворення РНК. Ці білки можуть діяти як специфічні реплікази або дерепресори, або як специфічні захисні чинники, захищаючі від деградації. Зв'язки між самовідтворюваними одиницями повинні утворювати накладений цикл — лише тоді система у цілому буде подібна до гіперциклу. Представником систем цього класу є посилений автокаталізатор. Наприклад, у випадку інфекції клітини РНК-містким фагом. Якщо фагова РНК проникає до бактеріальної клітини, то її генотипічна інформація транслюється апаратом клітини-господаря. Один з продуктів трансляції зв'язується з певними чинникам господаря із утворенням активного ферментнго комплексу.
Реплікація полінуклеотидів () каталізується поліпептидами які у свою чергу є продуктами трансляції полінуклеотидів. Гіперциклічний зв'язок встановлюється за допомогою динамічних зв'язків двох типів:


- Кожний полінуклеотид специфічно транслюється у поліпептид Для трансляції потрібна наявність відповідного апарату, який включає у себе принаймні декотрі з продуктів трансляції й використовує певний генетичний код.
- Полінуклеотиди й поліпептиди утворюють специфічні комплекси, які також проявляють каталітичну активність при синтезі полінуклеотидних копій. Поліпептиди можут бути специфічними репліказами або специфічними кофакторами білка із полімеразною активністю. Усі ці первинні білки забезпечують наявність принаймні двох функцій: специфічної реплікації й трансляції. Математично циклічна симетрія уводиться тим, що припускається утворення специфічного комплексу між ферментом та полінуклеотидом , причому




Кінетика синтезу полінуклеотидів описується рівнянням Міхаеліса-Ментен

Чотири нуклеозидтрифосфати та їх стехіометричні коефіцієнти позначаються відповідно через та [1]


Див. також
- Виникнення життя
- Хімічна еволюція
- Еволюція
- Модель квазівидів
- Модель сайзерів
Примітки
- M.Eigen, P.Shuster - The hypercycle.
Джерела
- Эйген М., Шустер П. Гиперцикл. Принципы самоорганизации макромолекул. М.: Мир, 1982. 270 с.
- Файстель Р. , Романовский Ю. М., Васильев В. А. Эволюция гиперциклов Эйгена, протекающих в коацерватах // Биофизика. 1980. Т.25. N.5. С. 882–887.
- Опарин А. И. О возникновении жизни на Земле. М.: Изд-во АН СССР, 1957.
- Опарин А. И. Жизнь, ее природа, происхождение и развитие. М.: Наука, 1968.
- Ратнер В. А., Шамин В. В. Сайзеры: моделирование фундаментальных особенностей молекулярно-биологической организации // Математические модели эволюционной генетики. Новосибирск: ИЦИГ, 1980. С. 66 — 126.
- Ратнер В. А., Шамин В. В. Сайзеры: моделирование фундаментальных особенностей молекулярно-биологической организации. Соответствие общих свойств и конструктивных особенностей коллективов макромолекул // Журн. общ. биологии. 1983. Т.44. N.1. С. 51-61.
