Трансляція (біологія)
Трансляція — синтез білків з амінокислот, що каталізується рибосомою на матриці матричної РНК (мРНК). Трансляція є однією зі стадій біосинтезу білків, а він, у свою чергу, — частина процесу експресії генів.
Трансляція відбувається в цитоплазмі, де знаходяться рибосоми клітини. Під час трансляції інформація, що міститься в мРНК, розшифровується згідно з правилами, відомими як генетичний код, та використовується для синтезу закодованої поліпептидної послідовності. Процес трансляції можна поділити на чотири фази: активацію, ініціацію, елонгацію та термінацію.
При активації відповідна амінокислота приєднується до відповідної транспортної РНК (тРНК). Хоча ця стадія часто розглядається окремо від трансляції, вона необхідна для її початку. Зв'язана з амінокислотою тРНК називається аміноацил-тРНК, або «зарядженою» тРНК. При ініціації мала субодиниця рибосоми зв'язується з 5'-кінцем мРНК за допомогою факторів ініціації (IF) та інших білків, що допомагають процесу. Елонгація відбувається, коли чергова аміноацил-тРНК використовується для збільшення поліпептидного ланцюжка. Термінація відбувається, коли рибосома зустрічає стоп-кодон (UAA, UAG або UGA), для якого не існує відповідної тРНК; при цьому відбувається звільнення поліпептидного ланцюжка.
Механізм трансляції
Загальні відомості
Для здійснення трансляції в клітинах усіх без винятку організмів існують спеціальні органели — рибосоми. Рибосоми є рибонуклеопротеїдними комплексами, побудованими з 2 субодиниць: великої і малої. Функція рибосом полягає в розпізнаванні тринуклеотидних кодонів мРНК, підбору відповідних ним амінокислот і приєднанні цих амінокислот до білкового ланцюжка, що росте. Рухаючись уздовж молекули мРНК, рибосома розпізнає кодон за кодоном і синтезує білок відповідно інформації, закладеної в молекулі мРНК.
Для розпізнавання амінокислот в клітині існують спеціальні «адаптери», молекули транспортної РНК (тРНК). Ці молекули, що мають форму конюшинового листа, мають ділянку (антикодон), комплементарну кодону мРНК, та іншу ділянку, до якої приєднується амінокислота, що відповідає цьому кодону. Приєднання амінокислот до тРНК здійснюється в екзоенергетичній реакції ферментами аміноацил-тРНК-синтетазами, а молекула, що отримується в результаті, називається аміноацил-тРНК. Таким чином, специфічність трансляції визначається взаємодією між кодоном мРНК і антикодоном тРНК, а також специфічністю аміноацил-тРНК-синтеназ, що приєднують амінокислоти строго до відповідних їм тРНК (наприклад, кодону GGU відповідатиме тРНК, що містить антикодон CCA, а до цієї тРНК приєднуватиметься тільки амінокислота гліцин).
Механізми трансляції прокаріотів (бактерій та архей) і еукаріотів істотно відрізняються, тому багато речовин, що пригнічують прокаріотичну трансляцію, в значно меншому ступені діють на трансляцію еукаріотичних організмів, що дозволяє використовувати їх у медичній практиці як антибактеріальні засоби, безпечні для організму ссавців.
Оскільки кожен кодон містить три нуклеотиди, один і той самий генетичний «текст» можна прочитати трьома різними способами (починаючи з першого, другого і третього нуклеотидів), тобто в трьох різних рамках зчитування. За деякими цікавими винятками, значущою є інформація, закодована тільки в одній рамці зчитування. З цієї причини украй важливим для синтезу білка рибосомою є її правильне позиціювання на стартовому AUG-кодоні — під час ініціації трансляції.
Ініціація. 1. Розпізнавання стартового кодону (AUG), супроводжується зв'язуванням тРНК аміноацилированої метіоніном (М) і збіркою рибосоми з великої і малою субодиниць.
Елонгація. 2. Розпізнавання поточного кодону відповідною йому аміноацил-тРНК (комплементарна взаємодія кодону мРНК і антикодону тРНК збільшена). 3. Приєднання амінокислоти, принесеної тРНК, до кінця поліпептідного ланцюжка, що росте. 4. Просування рибосоми уздовж матриці, що супроводжується вивільненням молекули тРНК. 5. Аміноацилювання молекули тРНК, що вивільнилася, відповідній їй аміноацил-тРНК-синтетазою. 6. Приєднання наступної молекули аміноацил-тРНК, аналогічно стадії (2). 7. Рух рибосоми молекулою мРНК до стоп-кодона (в даному випадку UAG).
Термінація. Розпізнавання рибосомою стоп-кодона супроводжується (8) від'єднанням новосинтезованого білка і в деяких випадках (9) дисоціацією рибосоми.
Ініціація
Синтез білка завжди починається з AUG-кодону, що також кодує метіонін. Цей кодон зазвичай називають стартовим або ініціаторним. Ініціація трансляції передбачає пізнавання рибосомою цього кодону і залучення ініціаторної аміноацил-тРНК. Для ініціації трансляції необхідна також наявність певних нуклеотидних послідовностей в районі стартового кодону. Існування послідовності, що відрізняє стартовий AUG від внутрішніх, абсолютно необхідне, оскільки інакше ініціація синтезу білка відбувалася б хаотично на всіх AUG-кодонах.
Процес ініціації забезпечується спеціальними білками — факторами ініціації (англ. initiation factors, скорочено IF).
Мала рибосомна субодиниця (30S) прокаріотів, якщо вона не залучена в цей час в трансляцію, існує в комплексі з факторами ініціації IF1, IF3 і, в деяких випадках, IF2:
- IF3, зв'язаний з 30S-субодиницею, запобігає асоціації з великою (50S) субодиницею рибосоми, тим самим зберігаючи її вільний стан до зв'язування з матричною РНК. Цей білок також бере участь в скріпленні мРНК і тРНК, а також IF2;
- IF2 взаємодіє з тРНК, а також володіє здатністю розщеплювати ГТФ;
- IF1 є, мабуть, необов'язковим фактором (у деяких видів він відсутній), який підвищує спорідненість малої субодиниці до IF2 і IF3.
Комплекс 30S-субодиниці з ініціаторними факторами здатний розпізнавати спеціальні послідовності мРНК, так звані ділянки зв'язування рибосоми (англ. ribosomt-binding site або RBS). Ці ділянки містять, по-перше, ініціаторний кодон AUG і, по-друге, спеціальну послідовність Шайн-Дальгарно, з якою комплементарно зв'язується рибосомна 16S РНК. Послідовність Шайн-Дальгарно служить для того, щоб відрізнити ініціаторний AUG від внутрішніх кодонів, що кодують метіонін. Після того, як 30S-субодиниця зв'язалася з мРНК, до неї притягується ініціаторна аміноацил-тРНК і IF2, якщо вони ще не були включені в комплекс. Потім приєднується 50S-субодиниця, відбувається гідроліз ГТФ і дисоціація факторів ініціації. Зібрана рибосома починає синтезувати поліпептидний ланцюжок.
Початкова стадія передбачає зв'язування малої рибосомної субодиниці (30S) з мРНК. Це може відбуватися двома способами: або спочатку до мРНК приєднується комплекс, що містить рибосомну субодиницю (1), а потім до нього притягується тРНК в комплексі з IF2 і ГТФ (2), або 30S-субодиниця спочатку зв'язується з тРНК, а вже потім сідає на мРНК (3). До комплексу, що утворився, приєднується велика (50S) рибосомна субодиниця (4), фактори ініціації від'єднуються від 30S-субодиниці, що супроводжується гідролізом ГТФ білком IF2 (5) і зібрана рибосома починає елонгувати ланцюжок (6).
У правому нижньому кутку дана схема ініціаторної ділянки прокаріотичної мРНК. Відмічені 5'- і 3'-кінці молекули. RBS — ділянка зв'язування рибосоми, SD — послідовність Шайн-Дальгарно, AUG — ініціаторний кодон.
Елонгація
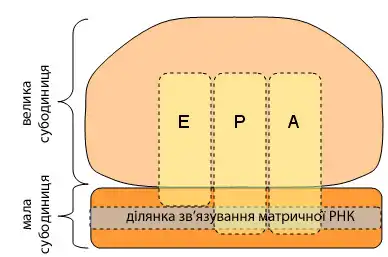
Елонгація поліпептидного ланцюжка полягає в додаванні нових амінокислот до карбоксильного (C-) кінця ланцюжка, що росте. Цей поліпептидний ланцюжок виходить з рибосоми через вихідний тунель у великий субодиниці.
Елонгація починається, коли метильована аміноацил-тРНК зв'язується з ділянкою P, що приводить до конформаційної зміни комплексу, яка відкриває ділянку A для зв'язування нової аміноацил-тРНК. Це зв'язування полегшується фактором елонгації Tu (EF-TU), малою ГТФазою. У цей момент ділянка P містить початок поліпепдидного ланцюжка, що синтезується, а ділянка A містить наступну амінокислоту, яка має бути додана до ланцюжку. Після цього поліпептид відділяється від тРНК в ділянці P і пептидний зв'язок формується між останньою амінокислотою поліпептида і амінокислотою, що все ще приєднана до тРНК в ділянці A. Цей процес, відомий як утворення пептидного зв'язку, каталізується рибозимом, пептидилтрансферазою, така активність властива до 23S-рРНК великої (50S) рибосомної субодиниці. Після утворення пептидного зв'язку ділянка A містить поліпептид, тоді як ділянка P містить незаряджену тРНК (тРНК без амінокислоти).
На кінцевій стадії елонгації рибосома переміщається на три нуклеотиди у напрямку до 3'-кінця мРНК. Через те, що тРНК зв'язані з мРНК за рахунок спаровування кодон-антикодон, тРНК рухається відносно рибосоми, рухаючи поліпептид з ділянки A у ділянку P, а незаряджена тРНК переміщається у ділянку виходу (ділянку E). Цей процес каталізується фактором елонгації G (EF-G).
Рибосома продовжує транслювати кодони, що залишилися, тому що нові аміноацил-тРНК зв'язуютьться з ділянкою A, поки рибосома не зустріне кодон зупинки на мРНК (UAA, UGA або UAG).
Термінація і переробка
Термінація відбувається, коли один з трьох стоп-кодонів переміщається в ділянку A. Ці кодони не мають відповідних тРНК. Натомість, їх визнають спеціальні білки — фактори термінації (англ. release factors, RF), а саме RF1 (що розпізнає стоп-кодони UAA і UAG) або RF2 (що розпізнає стоп-кодони UAA і UGA). Третій фактор звільнення RF-3 каталізує звільнення RF-1 і RF-2 в кінці процесу термінації. Ці фактори каталізують гідроліз ефірного зв'язку, що зв'язує тРНК з пептидом, та вивільнення недавно синтезованого білка з рибосоми.
Пост-термінаційний комплекс, сформований після термінації, складається з мРНК із стоп-кодоном в ділянці A рибосоми і тРНК. Крок переробки рибосоми відповідає за розбирання пост-трансляційного рибосомного комплексу. Як тільки протеїн, що синтезується, звільняється після термінації, фактори переробки рибосоми і фактор елонгації EF-G звільняють мРНК і тРНК з рибосоми і роз'єднують 70S-рибосоми на 30S- і 50S-субодиниці. IF-3 також допомагає процесу переробки, запобігаючи повторному зв'язуванню субодиниць за рахунок зв'язування із 30S субодиницею. Цей процес готує рибосому для повторення циклу трансляції.
Полісоми
Трансляція зазвичай здійснюється більш ніж однією рибосомою одночасно. Через відносно великий розмір рибосом вони можуть зв'язуватися з ділянками мРНК на відстані не менше 35 нуклеотидів. Кілька рибосом і молекула мРНК, по якій вони рухаються, називаються полісомою, або полірибосомою.
Кеп-залежна ініціація
За допомогою цього механізму транслюється переважне число еукаріотичних мРНК. Білки, що беруть участь в процесах ініціації трансляції у еукаріотів називають eIF (англ. eukaryotic Initiation Factors — еукаріотичні фактори ініціації). Крім факторів ініціації eIF1, eIF2 і eIF3, що зв'язуються з малою рибосомною субодиницею (40S), і за своїми функціями є приблизно аналогічними відповідним білкам прокаріотів, еукаріоти мають ще дві групи факторів ініціації: сімейство факторів, що зв'язують мРНК — eIF4, і сімейство факторів, що зв'язуються з великою (60S) субодиницею рибосоми, eIF5. Нижче приведений список основних факторів:
- eIF4A — РНК-геліказа, фермент, що розплітає вторинну структуру мРНК для того, щоб рибосома могла по ній рухатися;
- eIF4B — привертає фактор eIF4A до молекули мРНК;
- eIF4E — зв'язує кеп, 7-метилгуанін, розташований на 5'-кінці молекули мРНК;
- eIF4G — потрібний для організації компонентів, що беруть участь в ініціації трансляції, в єдиний комплекс. Містить ділянки скріплення eIF4B, eIF4E, рибосоми;
- eIF5 — потрібний для залучення великої субодиниці рибосоми.
На першому етапі ініціації трансляції мала субодиниця рибосоми в комплексі з факторами ініціації eIF4G, eIF4B, eIF4E й ініціаторній тРНК приєднується до 5'-кінця мРНК за рахунок здатності eIF4E зв'язувати кеп-структуру, а білка eIF3 — мРНК. Потім білок eIF4B привертає геліказу eIF4A, і та починає розплітати мРНК у напрямку до 3'-кінцю, що супроводжується витратами енергії у формі молекул АТФ. За рахунок роботи цього білка 40S-субодиниця звільняється від білків eIF4G і eIF4E, і в комплексі з факторами ініціації, що залишилися, рухається по мРНК до ініціаторного кодону AUG, де відбувається дисоціація факторів ініціації, що залишилися, і залучення 60S-субодиниці рибосоми за допомогою eIF5, після чого починається синтез поліпептидного ланцюжка.
Кеп-незалежна ініціація
Тоді як в більшості випадків еукаріотична трансляція вимагає наявності кепа на 5'-кінці мРНК, деякі вірусні і клітинні мРНК обходять кеп-залежний механізм за рахунок ініціації трансляції на певних послідовностях всередині молекули РНК.
Найкраще дослідженим (але далеко не єдиним) прикладом кеп-незалежної трансляції в еукаріотах є так звана «внутрішня ділянка входу рибосоми» (англ. Internal Ribosome Entry Site або IRES). На відміну від кеп-залежного механізму, кеп-незалежний механізм також не вимагає сканування рибосомою від 5'-кінця рибосоми до стартового кодону. Рибосома можу бути доставлена до стартової ділянки IRIS за допомогою ITAF'ів, (IRES trans-acting fantors), що дозволяють обходити необхідність сканування від 5'-кінця РНК.
Цей метод трансляції був знайдений відносно недавно і є необхідним за умовах, які вимагають трансляції певних мРНК в стресових умовах, коли загальна ефективність трансляції зменшена. Приклади включають фактори, що викликають апоптоз, імуноглобуліни, деякі фактори росту. Крім того, цим механізмом інколи користуються віруси[1].
Елонгація
Елонгація трансляції еукаріотів дуже подібна до елонгації трансляції прокаріотів. Основними факторами елонгації є такі:
- eEF-1, чиї α- і βγ-субодиниці відповідають прокаріотичним факторам EF-TU і EF-TS відповідно;
- eEF-2, що відповідає прокаріотичному фактору EF-G.
Термінація
У еукаріотів існує лише один фактор вивільнення, eRF, замість трьох факторів прокаріотів. Проте загалом процес термінації подібний до процесу термінації прокаріотів[2].
Трансляція вручну
Принцип
Для визначення поліпептидної послідовності можливо провести процес перекладення послідовності ДНК у послідовність амінокислот, так звану «трансляцію вручну» (для коротких послідовностей) або трансляцію на комп'ютері. На першому кроці необхідно перевести послідовність ДНК у послідовність РНК, замінюючи нуклеотиди A, T,C, G в ДНК на U, A,G, C в РНК, відповідно.
Потім РНК розбивається на тринуклеотидні послідовності (кодони). Відзначте, що існує 3 «рамки зчитування», або способи інтерпретації коду залежно від того, на якому нуклеотиді почнеться трансляція.
Нарешті, використовується так звана «таблиця трансляції» для переводу кожного кодону у відповідну амінокислоту. В результаті отримується первинна структура білка.
Проте білок також має структуру вищого порядку (вторинну, третинну і четвертинну) або конформацію, яка визначається наявністю гідрофобних та електростатичних взаємодій між частинами поліпептидного ланцюжка. Тоді як вторинна структура (структура «доменів» — коротких ділянок білка) ще може буди передбачена за допомогою послідовності, належна третинна структура зазвичай важка для визначення теоретично, і навіть в природі вибір третинної структури часто залежить від взаємодії з іншими білками.
Трансляція вручну в деяких випадках також не дає правильної амінокислотної послідовності природного білка, зокрема якщо кодується нестандартна амінокислота, наприклад селеноцистеїн, який кодується за допомогою звичайного стоп-кодону UGA та спеціального «шпилькоподібного» РНК-мотиву, так званого SECIS-елементу.
Таблиці трансляції
Для більшості еукаріотів зазвичай використовується стандартна таблиця трансляції, у якій кожній амінокислоті відповідає одна або кілька послідовностей ДНК: (див. NCBI Taxonomy).
AAs = FFLLSSSSYY**CC*WLLLLPPPPHHQQRRRRIIIMTTTTNNKKSSRRVVVVAAAADDEEGGGG Starts = ---M---------------M---------------M---------------------------- Base1 = TTTTTTTTTTTTTTTTCCCCCCCCCCCCCCCCAAAAAAAAAAAAAAAAGGGGGGGGGGGGGGGG Base2 = TTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGGTTTTCCCCAAAAGGGG Base3 = TCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAGTCAG
Не всі організми використовують однаковий генетичний код. Навіть працюючи із генетичними послідовностями звичайних еукаріотичних організмів, наприклад дріжджів, часто бажано використовувати альтернативні таблиці трансляції — а саме для трансліції мітохондріальних генів. Зараз група таксономії NCBI визначає наступні таблиці для послідовностей, що містить GenBank (таблиці див. NCBI Taxonomy):
1: Стандартний код 2: Мітохондріальний код хребетних 3: Мітохондріальний код дріжджів 4: Мітохондріальний код слизистих грибків, протозоїв і кишковопорожниних, та мікоплазми/спіроплазми 5: Мітохондріальний код безхребетних 6: Ядерний код джгутикових, Dasycladacea і Hexamita 9: Мітохондріальний код голошкірих та плоських червів 10: Ядерний код Euplotida 11: Бактеріальний код і пластидний код рослин 12: Альтернативний ядерний код дріжджів 13: Мітохондріальний код асцидій 14: Альтернативний мітохондріальний код плоских червів 15: Ядерний код Blepharisma 16: Мітохондріальний код Chlorophycea 21: Мітохондріальний код Trematoda 22: Мітохондріальний код Scenedesmus obliquus 23: Мітохондріальний код Thraustochytrium
Комп'ютерна трансляція
Існує багато комп'ютерних програм, здібних до трансляції послідовності ДНК/РНК у білкову послідовність. Проте небагато програм здатні видавати правильну послідовність у всіх «особливих» випадках, таких як використання альтернативних ініціаторних кодонів. Наприклад рідкісний код альтернативного кодону ініціації TTG використовується для метіоніну, коли використовується як ініціаторний кодон, і для лейцину в решті випадків.
Примітки
- Brett D. Keiper and Robert E. Rhoads (1997). Cap-independent translation initiation in Xenopus oocytes. Nuclear Acids Research 25 (2): 395–402.
- Termination of translation: interplay of mRNA, rRNAs and release factors?. Процитовано 28 листопада 2006.
Посилання
- ApE Комп'ютерна програма трансляції (Mac, Windows, Unix)
- ExPASy Translate Tool Комп'ютерна програма трансляції (webserver)
- Virtual Ribosome Комп'ютерна програма трансляції (webserver, cross-platform command-line)
- Pamela C Champe, Richard A Harvey and Denise R Ferrier (2005). Lippincott's Illustrated Reviews: Biochemistry (3rd ed.). Lippincott Williams & Wilkins. ISBN 0-7817-2265-9.
- David L. Nelson and Michael M. Cox (2005). Lehninger Principles of Biochemistry (4th ed.). W.H. Freeman. ISBN 0-7167-4339-6.
Джерела
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет". с. 221-274.
