Хімічна еволюція
Хімі́чна еволю́ція чи пребіоти́чна еволюція — етап, що передував появі життя[1][2][3], в ході якого органічні, пребіотичні речовини виникли з неорганічних молекул під впливом зовнішніх енергетичних і селекційних факторів і через розгортання процесів самоорганізації, властивих усім відносно складним системам, якими, безперечно, є всі молекули, що мають Карбон.
Також ці терміни позначають теорію виникнення і розвитку тих молекул, які мають важливе значення для утворення і розвитку живої речовини.
Загальна характеристика
Все, що відомо про хімізм речовини, дозволяє обмежити проблему хімічної еволюції рамками так званого «воднево-вуглецевого шовінізму», який постулює, що життя в нашому Всесвіті представлене в єдиному можливому варіанті: як «способі існування білкових тіл»[4], утвореному завдяки унікальному поєднанню полімеризаційних властивостей вуглецю і деполяризаційних рідко-фазних властивостей водного середовища, як спільно необхідних і/або достатніх (?) умов для виникнення і розвитку всіх відомих нам форм життя. При цьому мається на увазі, що принаймні в межах однієї сформованої біосфери може існувати тільки один, загальний для всіх живих істот даної біоти код спадковості, але поки залишається відкритим питання, чи існують інші біосфери поза Землею і чи можливі інші варіанти генетичного апарату.
Також невідомо, коли й де почалася хімічна еволюція. Можливі будь-які терміни після закінчення другого циклу зореутворення, що наступив після конденсації продуктів вибухів первинних наднових зірок, які постачають в міжзоряний простір важкі елементи (з атомною масою понад 26). Друге покоління зірок, вже з планетними системами, збагаченими важкими елементами, які необхідні для реалізації хімічної еволюції, з'явилося через 0,5-1,2 млрд років після Великого вибуху. При виконанні деяких цілком ймовірних умов, для запуску хімічної еволюції може бути придатна практично будь-яке середовище: глибини океанів, надра планет, їхні поверхні, протопланетні утворення та навіть хмари міжзоряного газу, що підтверджують повсюдні виявлення астрофізичними методами в космосі багатьох видів органічних речовин — альдегідів, спиртів, вуглеводів і навіть гліцину (амінокислота), які разом можуть бути матеріалом для хімічної еволюції, кінцевим результатом якої є виникнення життя.
Методологія дослідження хімічної еволюції (теорія)
Дослідження хімічної еволюції ускладнюється тим, що зараз знання про геохімічні умови древньої Землі не є достатньо повними.
Тому, крім геологічних, потрібні також астрономічні дані. Наприклад, умови на Венері та Марсі розглядають як близькі до тих, що були на Землі на різних етапах її еволюції.
Основні дані про хімічну еволюцію отримані через модельні експерименти, в ході яких вдалося отримати складні органічні молекули при імітації різних хімічних складів атмосфери, гідросфери та літосфери та кліматичних умов.
На основі наявних даних висунуто ряд гіпотез про конкретні механізми та безпосередні рушійні сили хімічної еволюції.
Абіогенез
Абіогенез — утворення органічних сполук, поширених в живій природі, поза організмом без участі ферментів.
У загальному розумінні абіогенез — виникнення живого з неживого, тобто вихідна гіпотеза сучасної теорії походження життя. У 1920-х роках академік Опарин припустив, що в розчинах високомолекулярних сполук можуть мимовільно утворюватися зони підвищеної концентрації, які відносно відокремлені від зовнішнього середовища і можуть підтримувати обмін з нею. Він назвав їх коацерватними краплями, або просто коацервати.
У 1953 Стенлі Міллером експериментально здійснено абіогенним синтез амінокислот та інших органічних речовин в умовах, що відтворюють умови первісної Землі.
Існує також теорія гіперциклів, згідно з якою перші прояви життя були пов'язані з т. зв. гіперциклами — комплексами складних каталітичних реакцій, в яких продукт кожної попередньої реакції є каталізатором для наступної.
У 2008 році американські біологи зробили важливий крок для розуміння початкових етапів зародження життя. Їм вдалося створити «протоклітини» з оболонкою з простих ліпідів і жирних кислот, здатну втягувати з навколишнього середовища нуклеотид-монофосфати, активовані імідазолом - «цеглинки», необхідні для синтезу ДНК[5]. У 2011 році японські вчені повідомили, що їм вдалося створити везикули з катіонною оболонкою і елементами ДНК всередині, здатну до поділу в результаті полімеразної ланцюгової реакції, реплікується ДНК.[6]
Цинковий світ
Гіпотези хімічної еволюції повинні пояснювати такі аспекти:
- Поява в Космосі або на Землі умов для автокаталітичного синтезу великих обсягів і значного різноманіття молекул Карбону, тобто — виникнення в абіогенних процесах речовин, необхідних і достатніх для початку хімічної еволюції.
- Поява з таких молекул щодо стійких замкнутих агрегатів, що дозволяють так ізолювати себе від навколишнього середовища, що з нею стає можливим виборчий обмін речовиною та енергією, тобто — виникнення деяких протоклітинних структур.
- Поява в таких агрегатах здатних до самозміни й до самореплікації хімічних інформаційних систем, тобто — виникнення елементарних одиниць спадкового коду.
- Поява взаємної залежності між властивостями білків і функціями ферментів з носіями інформації (РНК, ДНК), тобто — виникнення власне коду спадковості, як необхідної умови вже для біологічної еволюції.
Значний внесок у розв'язанні цих питань, серед інших, зробили такі вчені:
- Олександр Опарин: Коацервати.
- Гарольд Юрі та Стенлі Міллер в 1953: Виникнення простих біомолекул в симульованій древній атмосфері.
- Сідней Фокс: Мікросфери з протеноїдів.
- Томас Чек (університет Колорадо) і Сідней Алтман (університет Yale New Haven Connecticut) в 1981: автокаталітичний РНК-поділ: «Рібозими» об'єднують каталіз та інформацію в молекулі. Вони можуть вирізати себе з довшого ланцюга РНК і з'єднувати знову кінці, що залишилися.
- Волтер Гілберт (Гарвард університет, Кембридж) розробляє в 1986 ідею світу РНК.
- Гюнтер фон Кідровськи (Рур-університет Бохум) представляє в 1986 році першу самовідтворювану систему на основі ДНК, важливий внесок у розуміння функцій зростання самовідтворюваних систем
- Манфред Ейген (інститут Макса Планка факультет біофізичної хімії, Геттінген): Еволюція ансамблів молекул РНК. Гіперцикл.
- Юлій Ребека (Кембридж) створює штучну молекулу (Aminoadenosintriazidester), яка самовідтворюється в розчині хлороформу. Копії все ж ідентичні зразку, тому еволюція для цих молекул неможлива.
- Джон Корліс (Goddard центр космічних польотів - НАСА): Термальні джерела морів постачають енергію і хімікалії, які роблять можливими незалежну від космічного середовища хімічну еволюцію. Ще сьогодні вони є середовищем життя для початкових за багатьма ознаками археобактерій (Archaea).
- Гюнтер Вехтерсхойзер(Мюнхен) - гіпотеза залізо-сірчаного світу: перші самовідтворювані структури з обміном речовин виникли на поверхні піриту. Пірит (сульфід заліза) надавав для цього необхідну енергію. На кристалах піриту, які поступово зростали та розпадалися, ці системи могли рости та розмножуватися, та різні популяції протистояли в різних умовах середовища (умови відбору).
- А. Г. Cairns-Smith (університет Глазго) і Девід К. Мауерцалл (Rockefeller-Universität New York, Нью-Йорк) передбачають, що в глиняних мінералах є система, в якій відбувається хімічна еволюція, через що виникає багато різних, самовідтворюваних кристалів. Ці кристали притягують своїм електричним зарядом органічні молекули та каталізують синтез комплексних біомолекул, причому обсяг інформації кристалічних структур спочатку є матрицею. Ці органічні сполуки стають все складнішими доти, доки вони не зможуть розмножуватися без допомоги глиняних мінералів.
- Вольфганг Вайганд, Марк Дерр та ін. (Інститут Макса Планка факультет біогеохімії, Єна) показали у 2003, що сульфід заліза може каталізувати синтез аміаку з молекулярного азоту.
Уніфікована модель хімічної еволюції ще не розроблена, можливо, тому, що основні принципи ще не досліджені цілком.
Біомолекули
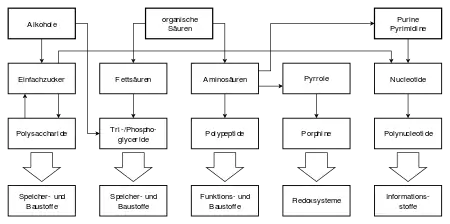
Пребіотичний синтез складних сполучень молекул може ділитися на три послідовні етапи:
- Виникнення простих органічних сполук (спиртів, кислот, гетероциклічних сполук : пуринів, піримідинів і піролів) з неорганічних матеріалів.
- Синтез складніших органічних сполук — «біомолекул» — представників найбільш поширених класів метаболітів, в тому числі й мономерів — структурних одиниць біополімерів (моносахаридів, амінокислот, жирних кислот, нуклеотидів) з простих органічних сполук.
- Виникнення складних біополімерів (полісахариди, білки, нуклеїнові кислоти) з основних структурних одиниць — мономерів.
Одним з питань є хімічний склад середовища, в якому здійснювався б пребіологічний синтез, в тому числі те, які неорганічні компоненти були б джерелами різних елементів, що входять до складу різних органічних сполук.
| C | H | O | N | S | P | |
|---|---|---|---|---|---|---|
| Вуглеводи | X | X | X | |||
| Жири | X | X | X | X | ||
| Фосфоліпіди | X | X | X | X | X | |
| Білки | X | X | X | X | X | |
| Нуклеотиди | X | X | X | X | X | |
| Порфірини | X | X | X | X |
| Відновлена форма | Окислена форма | |
|---|---|---|
| Карбон (C) | Метан (CH4), Монооксид вуглецю (чадний газ, CO) | Вуглекислий газ (CO 2) |
| Гідроген (H) | Водень (H2) | Вода (H2O) |
| Оксиген (O) | H2O | Кисень (O2) |
| Нітроген (N) | Аміак (NH 3) | Нітрати (NO3-) |
| Сульфур (S) | Сірководень (H 2 S) | Сульфати (SO42-) |
| Фосфор (P) | Фосфати (PO43-) |
<br>Всі гіпотези виходять з того, що, крім води та фосфатів, на початкових етапах історії Землі в атмосфері та гідросфері в достатній кількості були тільки відновлені форми, що відрізняються від звичайних в сучасний період хімічних сполук, оскільки стародавня атмосфера не містила молекулярного кисню.
Як джерело енергії, що ініціює синтез, в цей час могли виступати ультрафіолетове випромінювання Сонця, тепло вулканічних процесів, іонізаційне випромінювання радіоактивного розпаду та електричні розряди. Існують також теорії, в рамках яких джерелом необхідної для виникнення біомолекул енергії можуть служити окислювально-відновні процеси між вулканічними газами (відновник) і частково окислювальним сульфідними мінералами, наприклад, піритом (FeS2).
Цинковий світ
Розвиток земної атмосфери є однією з частин хімічної еволюції й до того ж важливим елементом історії клімату. Сьогодні її поділяють на чотири важливі етапи розвитку.
Спочатку відбувалося утворення хімічних елементів в космосі і поява з них Землі — приблизно 4,56 мільярда років тому. Ймовірно наша планета вже досить рано мала атмосферу з водню (H2) і гелію (He), яка, однак, знову розвіялася в космічний простір. Астрономи вважають, що через відносно високі температури та ефекти сонячного вітру на Землі та інших прилеглих до Сонця планетах могла залишитися тільки невелика кількість легких хімічних елементів (включаючи вуглець, азот і кисень ). Всі ці елементи, які складають сьогодні основну частину біосфери, були занесені, за цією теорією, ударами комет з більш зовнішніх ділянок сонячної системи лише через великий проміжок часу, коли протопланети трохи охололи. Протягом перших декількох мільйонів років після виникнення сонячної системи постійно повторювалися зіткнення з небесними тілами, викликані ними колізії знищували глобальними стерилізаціями утворені в цей час живі системи. Тому поява життя змогло розпочатися тільки після накопичення води за тривалий час хоча б в найглибших западинах.


З повільним охолодженням Землі, вулканічною діяльністю (виділення газів з надр Землі) і глобальним розподілом матеріалів комет, що впали, виникла друга атмосфера Землі. Швидше за все, вона складалася з водяної пари (H2O; до 80%), вуглекислого газу (CO2; до 20%), сірководню (H2S; до 7%), аміаку (NH3) і метану (CH4). Високий відсоток водяної пари пояснюється тим, що поверхня Землі була на той момент ще занадто гаряча для утворення морів. Перш за все з води, метану й аміаку в умовах молодої Землі могли утворитися невеликі органічні молекули (кислоти, спирти, амінокислоти), пізніше також органічні полімери (полісахариди, жири, поліпептиди), які були нестабільні в кислотній атмосфері.
Після охолодження атмосфери до температури, нижчої за кипіння води, настав дуже тривалий період випадання дощів, які й утворили океани. Насиченість інших газів атмосфери щодо водяної пари підвищилася. Інтенсивне ультрафіолетове випромінення зумовило фотохімічний розпад води, метану й аміаку, в результаті чого накопичилися вуглекислий газ і азот. Легкі гази — водень і гелій — вилетіли в космос, вуглекислий газ розчинявся у великих кількостях в океані, збільшуючи кислотність води. Значення pH сягнуло 4. Інертний і малорозчинний азот N2 накопичувався з часом і утворював близько 3,4 мільярда років тому основну складову атмосфери.
Випадання в осад прореагованого з іонами металів розчиненого вуглекислого газу (карбонати) і подальший розвиток живих істот, які асимілювали вуглекислий газ, призвело до зменшення CO2-концентрації та підвищення значення pH в водоймах.
Кисень O2 грає найважливішу роль в подальшому розвитку атмосфери. Він утворився з появою здатних до фотосинтезу живих істот, імовірно ціанобактерій (синьо-зелених водоростей) або їм подібних прокаріотів. Асиміляція ними вуглекислого газу призвела до подальшого зниження кислотності, концентрація кисню в атмосфері залишалася все-таки досить низькою. Причина — негайне використання розчиненого в океані кисню для окислення двовалентних іонів заліза та інших речовин, що окислюються з'єднань. Близько двох мільярдів років тому цей процес завершився, і кисень став поступово накопичуватися в атмосфері.
Дуже реакційноздатний кисень легко окислює сприйнятливі органічні біомолекули та стає в такий спосіб фактором відбору навколишнього середовища для ранніх організмів. Лише поодинокі анаероби змогли переміститися в вільні від кисню екологічні ніші, інша частина виробила ферменти (наприклад, каталази), які роблять кисень безпечним. У деяких мікроорганізмах з подібних ферментів розвинулися комплексні мембранні ферменти — кінцеві оксидази, які метаболічно використовували присутній кисень для накопичення енергії, необхідної для росту власної клітини — кінцева стадія окислення в аеробного ланцюга дихання. Залежно від організму є різні форми кінцевих оксидаз, наприклад, хінол-оксидаза або цитохром C — оксидаза, які розрізняються активними центрами, що містять іони міді й геми. Це дає підставу вважати, що вони відбулися різними паралельними шляхами розвитку. У багатьох випадках в одному організмі зустрічаються різні типи кінцевих оксидаз. Ці ферменти є останніми в ланцюзі послідовно залучених комплексів ферментів, які енергію зберігають окисно-відновних процесів перенесенням протонів або іонів натрію в формі трансмембранного електричного потенціалу. Останній іншим комплексом ферментів перетворюється знову в хімічну енергію у формі АТФ. Синтез АТФ та інших компонент ланцюга дихання в еволюційному світлі значно старіше кінцевих оксидаз, оскільки вони грали важливу роль вже в багатьох аеробних процесах обміну речовин (аеробне дихання, багато процесів бродіння, метаногенеза), а також при аноксигенному та оксигенному фотосинтезі.
Мільярд років тому вміст кисню в атмосфері переступило планку одного відсотка і через кілька мільйонів років був утворений озоновий шар. Сьогоднішній вміст кисню у 21% досягнуто лише 350 мільйонів років тому і зберігається з того часу стабільним.
Значення води для виникнення і збереження життя

Для життя, як ми його знаємо (або визначаємо), потрібна вода як універсальний розчинник. Вода має комплекс властивостей, які роблять життя можливим. Немає даних, що життя може виникати та існувати незалежно від води, і заведено вважати, що тільки наявність води в рідкій фазі (в певній області або на певній планеті), уможливлює виникнення життя.
Для синтезу складних молекул і дотримання граничних умов автокаталітичних реакцій між ними (що забезпечує можливість переходу від хімічної еволюції до еволюції живої речовини), важливими є такі характеристики води:
- Вода знаходиться в рідкому стані при температурі, в якій стабільні органічні молекули і їх синтез можливий тільки у водних розчинах.
- Вода необхідна як деполяризувальний розчинник для хімічних реакцій, бо вона робить можливим гомогенне перемішування, а маючи високу теплоємність, приймає теплоту, що виділяться, при реакціях і надає в розпорядження протони для каталізу.
- Висока питома теплоємність води, зосередженої в океані, і особливості характеристики спектрального поглинання водяної пари (при їх значущих частках в обсязі атмосфери) забезпечують стабілізацію глобальних коливань температури та осмотичних процесів (локально можуть виникати великі відмінності), що створює передумови до планетарно врівноваженого клімату, несприятливі зміни якого (для живої речовини), викликані як ентропійними процесами, так і зовнішніми несприятливими факторами, можуть бути в певних межах компенсовані життєдіяльністю самої біосфери, як цілого, що змінює інтенсивність випаровування води, і тим самим, планетарного альбедо і, що може бути ще більш важливо — змінюючи концентрацію в атмосфері водяної пари, головного по значенню парникового газу (див. гіпотезу Геї [7] британського вченого, консультанта NASA Джеймса Лаврока, а також конкурує з нею гіпотезу біотичної регуляції клімату [8] російського фізика В. Г. Горшкова).
- Вода і водяна пара поглинають шкідливе для макромолекул ультрафіолетове опромінення. Однак воно проникає через замерзлу воду (лід) до певної глибини.
- Вода, в якій розчинені речовини, наприклад, морська вода, утворює при виморожування області різних концентрацій речовин, які оточені мембранами льоду (див. гіпотеза морського льоду). Відокремленість від зовнішнього середовища і підвищена концентрація речовин вважаються необхідними для утворення біологічно активних молекул.
- Особливість термодинаміки води (при нагріванні від 0 до 3,98 ° С вода стискається) запобігає замерзанню водойм до дна - холодна вода, як менш щільна, залишається на поверхні і замерзає, а під льодом вода має позитивну температуру, що зберігало життя на Землі в епохи глобальних зледенінь.
Експерименти
Експеримент Міллера — Юрі
</br>Одними з найвідоміших гіпотез еволюції були думки російського дослідника А. І. Опарина у двадцятих роках XX століття та британського дослідника Дж. Голдейна. Теорія стверджувала, що умови на первісній Землі сприяли хімічним реакціям. З неорганічних сполук в атмосфері та морі повинні були синтезуватися складні органічні сполуки. Необхідна енергія постачалася дуже інтенсивним ультрафіолетовим опроміненням, яке могло безперешкодно проникати в атмосферу у зв'язку з малим вмістом в ній кисню та озону.
У 1953 році ця теорія була обґрунтована хіміками Стенлі Міллером і Гарольдом К. Юрі дуже хорошими результатами експерименту з «первинним бульйоном». Шляхом дослідів вони довели, що в середовищі, схожого на середовище з приблизною пребіотичними умовами, за допомогою припливу енергії ззовні (блискавки), з неорганічних сполук (вода, метан, аміак і водень) можуть виникнути амінокислоти та прості карбонові й жирні кислоти — одні з найважливіших будівельних елементів біомолекул (причому сучасні дослідження збереженого вмісту колб Міллера показало, що там містилося більшу кількість амінокислот, ніж зміг виявити Міллер[9]).
Пізніше в більшості випадків, в складніше побудованих дослідах з первинним бульйоном експериментатори змогли отримати як всі найважливіші будівельні елементи живих істот — амінокислоти, жири, цукру, нуклеотиди — так і складніші органічні сполуки — порфіни та ізопреноїди[10].
Як зауважив біохімік Роберт Шапіро, амінокислоти, синтезовані Міллером і Юрі — значно менш складні молекули, ніж нуклеотиди. Найпростіша з тих 20 амінокислот, що входять до складу природних білків, має всього два вуглецевих атомів, а 17 амінокислот з того ж набору — шість і більше. Амінокислоти та інші молекули, синтезовані Міллером і Юрі, містили не більше трьох атомів вуглецю. А нуклеотиди в процесі подібних експериментів вдалося отримати лише у 2009 р[11][12].
Хоча цим була показана можливість природного утворення органічних молекул, ці результати сьогодні іноді критикуються. В експерименті з первинним бульйоном виходили з того, що атмосфера на той період часу мала лужний характер, що відповідало науковим уявленням того часу. Сьогодні ж виходять зі слаболужного або навіть нейтрального характеру атмосфери, хоча питання ще не остаточно вирішене та обговорюються також локальні хімічні відхилення атмосферних умов, наприклад, в околицях вулканів. Пізнішими експериментами була доведена можливість появи органічних молекул і в цих умовах, навіть таких, що не утворилися при перших дослідах, але в значно менших кількостях. Це часто аргументує, що походження органічних молекул іншим шляхом, грало як мінімум додаткову роль. Приводяться також теорії походження органіки в околицях гідротермальних джерел серединно-океанічних хребтів.
Аргументом проти походження органічних молекул з первинного бульйону іноді наводять той факт, що під час досвіду виходить рацемат, тобто рівна суміш з L і D-форм амінокислот. Відповідно, повинен був існувати природний процес, в якому надавалася перевага певному варіанту хіральних молекул. Деякі астробіологи стверджують, що легше довести походження органічних сполук в космосі, оскільки, на їхню думку, фотохімічні процеси з циркулярно-поляризованим випромінюванням, наприклад, від пульсарів, в змозі знищити молекули тільки певного обертання. І дійсно, у знайдених в метеоритах хіральних органічних молекул переважали на 9% лівообертальні. Однак у 2001 році Alan Saghatelian[13] показав, що самовідтворювані пептидні системи теж в змозі ефективно відбирати молекули певного обертання в рацемічній суміші, що робить можливим і земне походження полімерів з певних оптичних ізомерів.
Подальші реакції
З проміжних продуктів альдегідів і синильної кислоти HCN, що з'являються під час проведення експерименту Міллера-Юрі, можна отримати при симульованих умовах Землі 4,5 млрд років тому подальші біомолекули. Таким чином Хуану Оро вдалося в 1961 році здійснити синтез аденіну:
| Едукти | продукти | ||
|---|---|---|---|
| 2-Формальдегід | HCN | H2O | Серин |
| 5-Формальдегід | Рибоза | ||
| 5 HCN | Аденін |
З рибози, аденіну і трифосфату виникає аденозинтрифосфат (АТФ), який використовується в організмах як універсальний енергоносій і будівельний елемент (як монофосфат) рибонуклеїнових кислот (РНК). Так само з похідних синильної кислоти (HCN), води (H2O) і аміаку (NH3) вдалося синтезувати гуанін і ксантин.[14]
Участь мінералів і гірських порід
- Кристалічні поверхні можуть служити матрицею для наростальних макромолекул. При цьому різні кристалічні поверхні можуть пов'язувати певні енантіомери молекул. L- та D-амінокислоти приєднуються до різних місць кристала кальциту.
- Аарон Качальський (Вайцман-інститут, Ізраїль) показав, що у водному розчині, що містить монтморилоніт (один з мінералів глин), можливий синтез білків з довжиною ланцюга в більше ніж 50 амінокислот з майже 100%-ним виходом.
- Іони металів можуть бути каталізаторами, донорами електронів або ж включатися в біомолекули.
- Мінерали глин у водних розчинах часто несуть поверхневий електричний заряд і можуть таким чином притягувати та утримувати протилежно заряджені органічні молекули.
- У мікропорожнинах гірських порід молекули органічних сполук захищені від ультрафіолетового випромінення.
Теорія Вехтерхойзера
Особливо інтенсивна форма сприяння мінералів і гірських порід пребіотичному синтезу органічних молекул повинна проходити на поверхні мінералів сульфіду заліза. Теорія Міллера-Юрі обмежується помилковим поясненням полімеризації мономерних складових біомолекули.
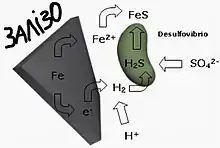

Альтернативний сценарій був з початку 1980-х років розроблений Гюнтером Вехтерхойзером. За цією теорією життя на Землі виникло на поверхні залізно-сірчаних мінералів, тобто сульфідів, які й сьогодні утворюються за допомогою геологічних процесів, а на молодій Землі повинні були зустрічатися набагато частіше. Ця теорія на противагу гіпотезі світу РНК, передбачає, що обмін речовин передував появі ферментів і генів. Як сприятливі місця пропонуються чорні курці на дні океанів, де високий тиск, висока температура, немає кисню і рясно представлені різні сполуки, які могли послужити будівельним матеріалом «цеглинок життя» або каталізатором в ланцюжку хімічних реакціях. Велика перевага цієї гіпотези перед попередніми в тому, що вперше утворення комплексних біомолекул пов'язано з постійним, надійним джерелом енергії. Енергія виділяється при відновленні частково окислених залізно-сірчаних мінералів, наприклад, піриту (FeS2), воднем (рівняння реакції: FeS2 + H2 ⇆ FeS + H2S), і цієї енергії досить для ендотермічного синтезу мономерних структурних елементів біомолекул і їх полімеризації:
Fe 2+ + FeS2 + H2 ⇆ 2 FeS + 2 H+; ΔH = -44,2 кДж/моль
Інші метали, так само як і залізо, теж утворюють нерозчинні сульфіди. На додаток до цього пірит і інші залізно-сірчані мінерали мають позитивно заряджену поверхню, на якій можуть розташовуватися, концентруватися і реагувати між собою переважно негативно заряджені біомолекули (органічні кислоти, фосфорні ефіри, тіоли). Необхідні для цього речовини (сірководень, монооксид вуглецю і солі двовалентного заліза) потрапляють з розчину на поверхню цього «залізо-сірчаного світу». Вехтерхойзер залучає до своєї теорії й сьогодні наявні основоположні механізми обміну речовин і виводить з них замкнутий в собі сценарій синтезу комплексних органічних молекул (органічні кислоти, амінокислоти, цукор, азотисті основи, жири) з простих неорганічних сполук, що знаходяться в вулканічних газах (NH3, H2, CO, СО2, CH4, H2S).
На противагу експерименту Міллера-Юрі не залучаються джерела енергії ззовні, у формі блискавки або ультрафіолетового випромінювання; крім того, перші щаблі синтезу при високих температурах і тисках протікають набагато швидше (наприклад, каталізуються ензимами хімічні реакції). При температурі підводних вулканів до 350°C виникнення життя є цілком ймовірним. Тільки пізніше при виникненні чутливих до високих температур каталізаторів (вітаміни, білки) еволюція повинна була відбуватися при нижчій температурі.
Сценарій Вехтерхойзера добре підходить до умов глибоководних гідротермальних джерел, оскільки наявний там перепад температури дозволяє подібний розподіл реакцій. Найдавніші з наявних нині мікроорганізмів — найбільш жаростійкі, граничний відомий температурний максимум для їх росту становить +122°C. Крім того, залізо-сірчані активні центри й сьогодні залучені в біохімічних процесах, що може вказувати на первинне участь Fe-S-мінералів в розвитку життя.
У цій теорії, однак, є суттєвий мінус, який полягає в тому, що фероплазма — мікроорганізм аеробний, а аеробні метаболізми з'явилися еволюційно набагато більш пізно, ніж анаеробні. (див. основну статтю)
Утворення макромолекул
Разом з гіпотезою «залізно-сірчаного» світу, в науковій спільноті розглядається ідея про «цинковий світ», запропонована біофізиком Арменом Мулкиджаняном[15][16]. Передбачається, що каталізаторами хімічних реакцій тут були сульфід цинку (ZnS) і сірководень (H2S), які отримували енергію з ультрафіолетового випромінювання.
Таким чином азот міг відновлюватися до аміаку[17]:

А вуглекислий газ до мурашиної кислоти:

Мурашина кислота та аміак, що утворюються на сульфіді цинку, при підсиханні води випадають в осад у вигляді форміату амонію HCOONH4. Ця сіль при прогріванні в сухому вигляді втрачає воду і перетворюється в формамід CHONH2. Формамід може бути сировиною для синтезу азотистих основ РНК[18][19]. Крім того, завдяки високій температурі кипіння (218°C при нормальному тиску) він в достатній кількості накопичується в висихних калюжах, і в середовищі формаміду, на відміну від водного середовища, нуклеотиди легко з'єднуються в РНК і не розпадаються.
Окрім того, сульфід цинку може приймати світлове збудження з нуклеотидів, РНК та інших молекул. Це захищає РНК від УФ-розщеплення, і водночас накопичення РНК на поверхні мінералу дозволяє збирати більше світла і прискорює «фотосинтез» в цьому місці.
Так само на користь «цинкового світу» свідчить той факт, що у відомих структурах РНК залізо абсолютно відсутнє, а цинк зустрічається частіше за інші перехідні метали. На другому місці — марганець, який поводить себе подібно цинку. Багато рибозимів вимагають присутності іонів металів для здійснення каталітичної активності. Серед таких металів найпоширеніші — магній, цинк і марганець, а залізо ніколи не зустрічається. [20]
Таким чином, найбільш сприятливим місцем для створення життя є не чорні курці, а грязьові вулкани, оскільки вони мають [21] [22] :
- середовище, збагачене калієм, фосфором та необхідними мікроелементами;
- місце перебування з влаштованим джерелом тепла, з практично постійними умовами незалежно від капризів погоди;
- пористі мінеральні осади, котрі працюють як каталізаторів і надають величезну кількість роздільних мікровідділів для перебування доклітинних форм життя;
- калюжі, що випаровуються та у котрих можуть накопичуватися органічні речовини й завдяки високій концентрації солей і формаміду може йти утворення ланцюжків РНК та білків;
- кілька різних механізмів отримання органічних речовин з атмосферного CO2 та азоту;
- освітлювану Сонцем поверхню, на котрій йдуть фотохімічні реакції, і зовсім поруч з нею — захищені від ультрафіолету пори в осадах;
- підігрів пор з однієї сторони й тріщини, у котрих відбувається накопичення нуклеотидів та РНК до високих концентрацій.
- відбір самих УФ-стійких азотистих основ;
- відбір азотистих основ, схильних утворювати комплементарні пари;
- відбір нуклеотидів одної хіральності з суміші правих і лівих нуклеотидів (т. зв. рацемічній суміші), тому що суміш правих і лівих нуклеотидів у ланцюжку порушує стекінг-взаємодію;
- відбір довгих молекул РНК у порівнянні з коротшими, тому що у довгих ланцюжках стекінг-взаємодія збільшує стійкість до ультрафіолету;
- відбір молекул РНК, котрі містили двоспіральні ділянки (шпильки), серед молекул з #випадковий послідовностями, тому що у їх більше нуклеотидів входять у склад комплементарних пар.
Ніякі інші місцеперебування не володіють відразу всіма цими перевагами. Наприклад, в «чорних курців» немає збагачення калієм і фосфором, немає ультрафіолету і немає накопичення речовин у випаровуваних калюжах. Тому грязьові котли наземних геотермальних полів на сьогодні є найімовірнішим місцем появи життя.
«Цинкову» теорію підтверджує ще той факт, що іони заліза легко розщеплюють РНК, зв'язуючись з 2 'і 3' гідроксильних груп рибози. Тому клітини зберігають надлишок заліза в пов'язаному вигляді, разом з білком феритином. У той час як іони цинку РНК молекулу не руйнують.
Утворення пребіотичних структур (попередників клітин)
Біомакромолекули — це білки й нуклеїнові кислоти. Збільшення молекулярних ланцюгів (полімеризація) потребує енергії та відбувається з виділенням води (конденсація). При розщепленні макромолекул (гідроліз) енергія виділяється. Оскільки хімічна рівновага настільки сильно зміщена в бік мономерів, що реакція протікає термодинамічно необоротно в бік гідролізу полімерів, синтез полімерів неможливий без постійного надходження енергії. Навіть за допомогою теоретичної підтримки випаровуванням води, додаванням солей (зв'язують воду) або розпадом продуктів, рівновага зміщується лише незначно. В результаті виникнення життя дуже ймовірно пов'язане з надійним джерелом енергії, яке використовувалося б для полімеризації.
[Мономери]n + H2O n Мономери + Тепло,


але
Енергія + мономери [Мономери]n + H2O.

Джерелом енергії в біохімії використовується найчастіше АТФ, для утворення якого необхідні вже створені ферменти. В умовах молодої Землі можливо постачання енергією синтезу полімерів гідролітичним розщепленням поліфосфатів, що використовується деякими ферментами замість розщеплення АТФ і сьогодні. Але малоймовірно, що поліфосфати вистачало, оскільки вони могли спонтанно утворюватися, при випаровуванні фосфатовмісних розчинах, але також відносно швидко гідролізувати, розчиняючись у воді. Схожі процеси могли відбуватися на узбережжі при регулярному припливі й відпливі. Але в цьому випадку всі залежні процеси від води постійно переривалися б, що занадто сповільнило б синтез комплексних сполук. Тому звернемося до зовсім іншої системи, в якій відбувається як синтез мономерних складових, так і залежне від постійного джерела енергії освіту полімерів, — до анаеробних окислювально-відновних реакцій з сульфідами металів.
Рівновага синтезу полімерів зміщується в бік утворення полімерів з підвищенням концентрації мономерів та зневодненням продуктів реакції. Умовою для цього є обмеження середовища реакції, яка має лише обмежений обмін речовин із зовнішнім середовищем. Вважалося, що подібні процеси протікають в маленьких ставках з високим випаровуванням, що спирається ще на ідеї праці Ч. Дарвіна. Сьогодні розглядаються вулканічні регіони океанів з осілими від гідротермальних джерел сульфідами металів як цілком ймовірне місце для розвитку схожих перетворень.
Інші варіанти розв'язання проблеми мають сильні обмеження і складно зіставляються з умовами ранньої Землі. Переважно потрібно для одного або декількох етапів виключення води, що дуже легко здійснити в лабораторії, але не в розглянутий час на землі. Одна з подібних систем — це полімеризація карбаміду (RN=C=NR) або диціану (N≡C-C≡N) в безводному середовищі. При цьому конденсація вихідних складових протікає паралельно реакції карбаміду, при якій виділяється необхідна енергія:
Енергія + [H-X-OH] + [H-X-OH] [X-X] + H2O (HX-OH = мономер, наприклад, амінокислота або рибоза)

[R-N = C = N-R] + H2O [R-NH-CO-NH-R] + Енергія (якщо R = H виникає сечовина)
При ультрафіолетовому випромінюванні з синильної кислоти утворюється диціан, проте у трясовині, що всихає, летка молекула швидко випарувалася б. Якщо суха суміш амінокислот нагрівається до 130°C кілька годин, то утворюються схожі на білок макромолекули. При наявності поліфосфатів досить 60°C. Ці умови можуть утворитися, якщо вода з розчиненими амінокислотами вступає в контакт з гарячим вулканічним попелом.
Якщо нагріти суміш нуклеотиду в присутності поліфосфатів до 55°C, то, хоча і виникають полінуклеотиди, але все ж з'єднання відбувається швидше шляхом 5'- і 2'-C-атомів рибози, оскільки воно протікає легше, ніж у всіх організмах з 5'-3'-зв'язками. З обох типів полінуклеотидів утворюються подвійні ланцюги (порівняйте з будовою ДНК). Зрозуміло, 5'-3'-подвійні ланцюги стабільніше, ніж 5'-2'.
Якщо на 2' атомі вуглецю рибози відсутня гідроксильна група, виходить дезоксорибоза. Тепер можуть утворюватися типові для ДНК 5'-3' зв'язки.
Світ РНК
Клітини підтримують свої функції, утворюючи окремі середовища для хімічних реакцій з метою поділу процесів обміну речовин і виключення небажаних реакцій. При цьому можуть створюватися різниця концентрацій.
Коацервати
Відомо, що з підвищенням концентрації багато органічних сполук, молекули яких містять як гідрофільні, так і гідрофобні ділянки, здатні у водних розчинах до міцелоутворення, тобто виділенню мікрокапель органічної фази. Міцелоутворення спостерігається також при висолюванні, тобто при збільшенні концентрації солей в колоїдних розчинах біополімерів-поліелектролітів, при цьому виділяються мікрокраплі діаметром 1-500 мкм, містять біополімери у високій концентрації.
Олександр Іванович Опарин (1894—1980) головним чином вивчав можливості обміну речовин у коацерватах (від латинського coacervatio — згусток або купа) — модельних міцелах. Їм було показано, що обмежені середовища з простим обміном речовин можуть виникнути виключно самоорганізацією, за умови присутності каталізаторів зі специфічними властивостями. Оскільки використані субстанції входять до складу, які живуть сьогодні організмів, Опаринські коацервати слід розглядати не як прабатьків клітин, а як модель виникнення таких прабатьків.
Опарин досліджував суміші з білків (гистон і альбумін), білків і вуглеводів (гистон і гуміарабік) і білків та полінуклеотидів (гистон або клупеїн і ДНК або РНК).
Якщо крапельки з гистона і гуміарабіку містять фермент фосфорилазу, то вони можуть поглинути з навколишнього середовища глюкозу-1-фосфат, створити з неї крохмаль і зберегти. Звільнений фосфат дифундує назовні. Капелька збільшується з накопиченням крохмалю, поки не розпадається на дрібні крапельки, які теж можуть містити фосфорилазу, зрозуміло, менше, ніж вихідна крапелька. Разом з тим сповільнюється обмін речовин. Отже, стає очевидним, що для збереження якостей клітини необхідна регенерація ферментів після її ділення.
Якщо до суспензії додати фермент амілазу, що розкладає крохмаль, виходять коацервати з двоступеневим обміном речовин: 1) поглинання глюкози-1-фосфату → синтез крохмалю; 2) вивід фосфату → розщеплення крохмалю і вивід мальтози.
Мікросфери
У 1970 році Сідней Фокс зміг підтвердити, що з подібних білку продуктів, які виникають при нагріванні сухих сумішей амінокислоти, а також самосполученням можуть виникнути зростальні крапельки, так звані мікросфери. Вони відмежовуються від навколишнього середовища мембраною зворотного осмосу і поглинають інший протеїноподібний матеріал з середовища. Внаслідок чого вони ростуть і розділяються знову на більш дрібні крапельки. На підставі цього Фокс прийшов до висновку, що ці системи мають ферментні якості, розкладають глюкозу або поводяться як естераза або пероксидаза, без додавання ферментів ззовні.
«Чорні курці»
Вперше гіпотеза світу РНК була висунута в 1986 році Уолтером Гільбертом і показала, що молекули РНК були попередниками організмів.
Гіпотеза відштовхується від здатності РНК до зберігання, передачі, і розмноженню генетичної інформації, а також від її здатності каталізувати реакції як рибозими. У еволюційному середовищі молекули РНК, які множать переважно себе, зустрічалися б частіше за інших.
Вихідним пунктом є прості самовідтворюються молекули РНК. Деякі з них мають здатність каталізувати синтез білків, які, своєю чергою, самі каталізують синтез РНК і свій власний синтез (розвиток трансляції). Деякі молекули РНК з'єднуються в подвійну РНК-спіраль, вони розвиваються в молекули ДНК і носії спадкової інформації (розвиток транскрипції).
Основою служать певні РНК-молекули, які можуть копіювати будь-які РНК-зразки, включаючи себе самих. Дженніфер А. Doudna і Джек В. Szostak використовували як зразок для розвитку цього типу РНК, що здійснює розрізання і сплайсинг самої себе інтрон Прокаріотних одноклітинного організму Tetrahymena thermophila. Це підтверджує те, що в рибосомах власне РНК є каталітичними молекулами та таким чином РНК каталізує синтез білка. Однак обмеження полягають у тому, щоб при самовідтворюваних РНК не моно-, а олігонуклеотиди є складовими ланками, а також необхідні допоміжні речовини.
У 2001 році відкрито, що важливими каталітичними центрами рибосом є РНК, а не, як раніше вважали, білки. Це показує, що каталітична функція РНК, як вона передбачалася в гіпотезі світу-РНК, використовується сьогодні живими істотами.
Оскільки рибосоми вважаються дуже початковими клітинними органоїдами, це відкриття вважається важливим внеском в обґрунтування гіпотези світу-РНК. Однак сьогодні не можна стверджувати, що молекули РНК можуть з амінокислот синтезувати білки, оскільки процесивної РНК-полімерази експериментально досі отримано не було.
У зв'язку з цим нуклеопротеїни (комплекси нуклеїнових кислот з білками) також можуть бути попередниками РНК.
Іншим попередником РНК могли бути поліциклічні ароматичні вуглеводні. Гіпотеза світу поліароматичних вуглеводнів намагається відповісти на питання, як виникли перші РНК, пропонуючи варіант хімічної еволюції від поліциклічних ароматичних вуглеводнів до РНК-подібних ланцюжків.
Земля весь час свого існування піддається бомбардуванню кометами та метеоритами. Особливо інтенсивними вони були відразу після утворення планети. У деяких метеоритах були виявлені прості органічні сполуки, серед інших амінокислоти. Одним з можливих доказів походження амінокислот в космосі могла б бути однакова хіральність органічних сполук (L-амінокислот і D-цукрів) — у метеоритних амінокислот доведено переважання L-типу до 9%[23]. Але цей розподіл може бути пояснений і дією неорганічних жирових каталізаторів. За допомогою дослідів, при яких симулювалися умови космосу, доведено, що основні органічні сполуки, особливо амінокислоти, можуть утворюватися і в таких умовах (Бернштейн, М. П., et al. 2002. «Racemic amino acids from the ultraviolet photolysis of interstellar ice analogues» Nature 416, 401 і Munoz Caro, G., et al. 2002. «Amino acids from ultraviolet irradiation of interstellar ice analogues» Nature 416, 403).
Умови середовища в цих гідротермальних джерелах, можливо, сприяли хемоавтотрофному виникненню життя. Ця теорія відстоюється дослідниками Вільямом Мартіном і Міхаелем Русселем[24].
Див. також
Примітки
- Futuyma D. J. Evolution. — Sunderland : Sinauer Associates, 2005. — P. 92—94. — ISBN 0-878-93187-2.
- Ridley M. Evolution. — 3rd ed. — Wiley-Blackwell, 2004. — P. 529—531. — ISBN 978-1-4051-0345-9.
- Rauchfuss, Horst. Chemical Evolution and the Origin of Life. — Springer, 2008. — ISBN 978-3-540-78822-5.
- Энгельс Ф. Анти-Дюринг; Маркс К. и Энгельс Ф., Сочинения, 2‑е изд., т. 20, с. 82.
- Искусственные протоклетки синтезируют ДНК без помощи ферментов
- С чего начиналось размножение на Земле. Архів оригіналу за 26 січня 2012. Процитовано 2 жовтня 2012.
- Элементы: Гипотеза Геи
- Биотическая регуляция: Ваши вопросы
- «Получены новые результаты старого эксперимента Стэнли Миллера»
- Эксперимент Миллера — Юри. lektsii.com. Процитовано 22 березня 2020.
- Matthew W. Powner, Béatrice Gerland, John D. Sutherland. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions // Nature. 2009. V. 459. P. 239—242.
- Химики преодолели главное препятствие на пути к абиогенному синтезу РНК • Новости науки. «Элементы» (рос.). Процитовано 8 листопада 2020.
- A. Saghatelian et al., 2001, A chiroselective peptide replicator, Nature, 409, 797—801
- Руттен М.Г. Эксперименты Оро с нагреваемой водной средой.
- Мулкиджанян Армен Яковлевич. On the origin of life in the Zinc world: I. Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth.
- Мулкиджанян Армен Яковлевич. On the origin of life in the zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth.
- Михаил Никитин. «Цинковый мир».
- S. D. Senanayake and H. Idriss. Photocatalysis and the origin of life: Synthesis of nucleoside bases from formamide on TiO2(001) single surfaces.
- Martin Ferus, Fabio Pietrucci, Antonino Marco Saitta, Antonín Knížek, Petr Kubelík, Ondřej Ivanek, Violetta Shestivska, and Svatopluk Civiš. Formation of nucleobases in a Miller–Urey reducing atmosphere.
- Михаил Никитин. Как сделать выбор между «цинковым» и «железосерным» мирами?.
- Михаил Никитин. Разные ответы на вопрос жизни.
- Михаил Никитин. Происхождение жизни. От туманности до клетки. — 2016.
- J.R. Cronin, S. Pizzarello, (2002) Enantiomeric Excesses in Meteoritic Amino Acids Science, 275, pp. 951—955
- M. Martin, M. J. Russell, (2003). On the origins of cells : a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleared cells. Philos. Trans. R. Soc. London Ser. B 358, 59-85
