Динорфіни
Динорфіни — клас опіоїдних пептидів, що утворюються з білка-попередника продинорфіну. Коли продінорфін розщеплюється під впливом пропротеїнової конвертази 2 (РС2), виділяються велика кількість активних пептидів: динорфін А, динорфін В та α / β-нео-ендорфін. Деполяризація нейрона, що містить продинорфін, стимулює процесинг PC2, яка відбувається в синаптичних пухирцях у пресинаптичному кінці. Іноді продинорфін переробляється не повністю, що призводить до вивільнення «великого динорфіну». «Великим динорфіном» є 32-амінокислотна молекула, що складається як з динорфіну А, так і з динорфіну В.[1]
| Динорфін | |
|---|---|
| Ідентифікатори | |
| Символ | PDYN |
| Інші ідентифікатори | |
| Інша інформація | |
Динорфін А, динорфін В та великий динорфін містять значну частку основних залишків амінокислот, зокрема лізину та аргініну (відповідно 29,4%, 23,1% та 31,2% залишків), а також гідрофобні залишки (41,2% , 30,8% та 34,4% відповідно). [2] Хоча динорфіни поширені в ЦНС, найвищі концентрації були виявлені в нейрогіпофізі[3], гіпоталамусі, довгастому мозку, мості, середньому мозку та спинному. [4] Динорфіни зберігаються у велитенських (діаметром 80-120 нм) щільних везикулах, які значно більші за нейромедіаторні. Ці великі пухирці відрізняються від синаптичних тим, що для їхнього вивільнення у синаптичну щілину необхідний більш інтенсивний та тривалий подразник. [5]
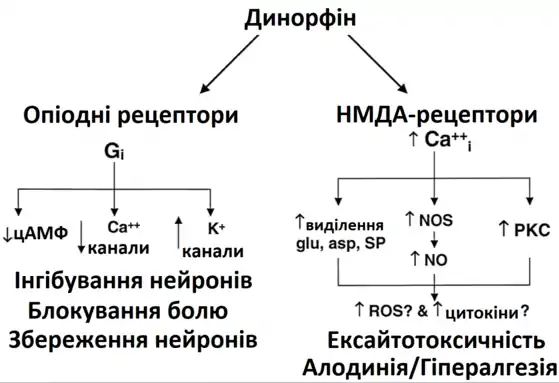
Перші ідеї про функції динорфінів сформували у 1979 Гольдштейн та ін. [6] у роботі з опіоїдними пептидами. Група виявила ендогенний опіоїдний пептид у гіпофізі свині, який було складно виділити. Після секвенування перших 13 амінокислот пептиду, вони створили синтетичну речовину зі схожими властивостями. Гольдштейн та ін. нанесли синтетичний пептид на поздовжній м’яз клубової кишки та виявили надзвичайно потужну опіоїдну дію. Таким чином, для опису дії пептиду, його назвали динорфіном (від грецького dynamis = сила). [6]
Динорфіни здійснюють ефект в основному за допомогою к-опіоїдного рецептора (КОР), зв'язаного з G-білками. Визначено два підтипи КОР: К1 та К2. [1] Хоча КОР є основним рецептором всіх динорфінів, ці пептиди також мають певну спорідненість до μ-опіоїдного рецептора (МОР), δ-опіоїдного рецептора (ДОР) та NMDA-рецептора. [5][7] Різні динорфіни демонструють неоднакову рецепторну селективність та силу зв’язування. Великий динорфін та динорфін А мають однакову селективність до КОР людини, але до МОР та ДОК саме великий динорфін має більшу селективність. Він також краще зв’язується із КОР, ніж динорфін А. Проте великий динорфін та динорфін А мають вищу селективність та силу зв’язування (визначається фактично значенням константи дисоціації комплекса агоніст-рецептор), ніж динорфін В. [8]
Синтез
Динорфіни утворюються в різних відділах нервової системи, включаючи гіпоталамус, нейрогіпофізі[3], стріатум, гіпокамп та спинний мозок. Схеми експресії генів з Allen Brain Atlas у мишей, макак та людей можна побачити тут.
Динорфіни мають багато різних фізіологічних ефектів, залежно від місця його синтезу.
- Наприклад, динорфін, який виробляється в магноцелюлярних вазопресинових нейронах супраоптичного ядра, важливий для формування електричної активності. Динорфін, що продукується в магноцелюлярних окситоцинових нейронах, є інгібітором негативного зворотного зв'язку секреції окситоцину.
- Динорфін, що виробляється в дугоподібному ядрі та в орексинових нейронах бічного гіпоталамуса, впливає на регуляцію апетиту.
Знеболення
Відомо, що динорфін є інгібітором больової чутливості. Також виявлено, що введення динорфіну в субарахноїдальний простір спинного мозку щурів викликало дозозалежне знеболення, яке визначалось затримкою руху хвоста. Аналгезію частково усунув антагоніст опіоїдів налоксон.[9]
Хан і Сі виявили, що динорфін в 6-10 разів потужніший за морфін.[9] Крім того, толерантність до морфію не зменшила індукований динорфіном знеболюючий ефект.[9] Рен та ін. продемонстрував певні проблеми, пов'язані із спричиненим динорфіном знеболюванням. Автори виявили, що поєднання субанальгетичних рівнів морфію та динорфіну А1-13 (версія динорфіну А, що містить лише перші 13 амінокислот пептиду) у спинному мозку щурів мала адитивні ефекти. Однак, коли динорфін А1-13 вводили у внутрішньоцеребровентрикулярну (ICV) область мозку, він мав антагоністичний ефект на створену морфієм аналгезію.[10]
Дослідження Lai та співавт. виявили, що динорфін також може стимулювати біль.[11] Група вчених знайшла, що він діє на рецептор брадикініну. Тирозиновий N-кінець динорфіну А необхідний для активації рецепторів, таких як К-опіодних рецепторів (КОР), але не потрібний для зв'язування з рецепторами брадикініну.[11] Лай та ін. вивчали ефекти динорфіну А2-13, який не містив тирозиновий N-кінець. На основі результатів досліджень динорфіну А2-13 автори запропонували механізм, за допомогою якого динорфін А активує брадикінінові рецептори і, таким чином, стимулює больову реакцію.[11]
Відповідно до цього механізму, динорфін активує рецептори брадикініну, що впускає іони кальцію в клітину через потенціал-чутливі канали. Блокування брадикінінових рецепторів у поперековому відділі спинного мозку зменшило стійкий біль.[11]
Свенсон та співавт. описав інший можливий механізм, за допомогою якого динорфін може активувати біль через спинний мозок.[12] Автори встановили, що введення укороченого динорфіну А2-17, який не зв'язується з опіоїдними рецепторами, спричинює збільшення кількості фосфорильованої р38 мітоген-активованої протеїнкінази (МАПК) у мікроглії заднього рогу спинного мозку. Активований р38 раніше був пов'язаний із NMDA-спричиненним виділенням простагландинів, що викликають біль.[13] Таким чином, динорфін також може викликати біль у спинному мозку через неопіоїдний шлях p38.
Інші дослідження виявили роль динорфіну та стимуляції Каппа-опіоїдних рецепторів за невропатичного болю.[14] Ця ж група також показала, що система динорфін-KOР опосередковує проліферацію астроцитів шляхом активації p38 MAПK, необхідного для ефекту невропатичного болю на знеболюючі відповіді.[15] У сукупності ці звіти припускають, що динорфін може викликати множинні ефекти як на Каппа-опіоїдні шляхи, так і на неопіоїдні для модуляції анальгетичних реакцій.
Звикання
Залежність від кокаїну є наслідком складних молекулярних змін у мозку після багаторазового впливу цієї речовини. Показано, що динорфіни є важливою частиною цього процесу. Хоча одноразовий вплив кокаїну не впливає на рівень динорфіну в мозку, повторний вплив збільшує концентрацію динорфіну в смугастому тілі та чорній речовині у щурів.[16]
Один із запропонованих молекулярних механізмів підвищення рівня динорфіну включає регуляцію транскрипції за допомогою CREB (транскрипційний фактор CRE; cAMP response element-binding protein). Відповідно до моделі, запропонованої Карлезоном та ін., вживання кокаїну збільшує експресію цАМФ та цАМФ-залежної протеїнкінази (протеїнкіназа А, ПКА). ПKA призводить до активації CREB, що збільшує експресію динорфіну в прилеглому ядрі та дорсальному смугастому тілі, тобто в тих областях головного мозку, що відповідають за звикання. Динорфін зменшує вивільнення дофаміну, зв'язуючись з К-опіодними рецепторами (КОР) на нервових закінченнях дофамінових волокон.[17]
Останні результати
Останні дослідження виявляють, що стреси та наркотична залежність стимулюють фактори комплексу динорфін/КОР. Тобто як і стрес, так і певні наркотики (наприклад, кокаїн), стимулюють надмірну експресію гену динорфіну, який зв'язується з K-опіоїдним рецептором (КОР). Це може бути наслідком дисфорії або депресії, перенесеними, наприклад, під час критичної стресової ситуації або під час симптомів абстиненції. Дані факти підтверджують те, що зв'язок динорфін/КОР бере участь у стресових поведінкових реакціях (дисфорії та депресії).[18]
Стрес та депресія
Стрес викликає вивільнення динорфінів через кортикотропний рилізінг-фактор та подальшу активацію КОР в різних областях мозку, які беруть участь в обробці емоцій: заднє ядро шва, гіпоталамус, вентральна тегментальна зона; це спричиняє дисфорію. Проте, ефекти можуть бути нівельовані, якщо заблокувати рецептори KOР за допомогою антидепресантів.[19]
Клінічне значення
Як правило, похідні динорфіну не мають клінічного застосування через коротку тривалість дії.[20]
Примітки
- Nyberg, Fred; Hallberg, Mathias (2007). Neuropeptides in hyperthermia. Progress in Brain Research 162. с. 277–293. ISSN 0079-6123. PMID 17645924. doi:10.1016/S0079-6123(06)62014-1. Процитовано 7 грудня 2020.
- Marinova, Zoya; Vukojevic, Vladana; Surcheva, Slavina; Yakovleva, Tatiana; Cebers, Gvido; Pasikova, Natalia; Usynin, Ivan; Hugonin, Loïc та ін. (15 липня 2005). Translocation of dynorphin neuropeptides across the plasma membrane. A putative mechanism of signal transmission. The Journal of Biological Chemistry 280 (28). с. 26360–26370. ISSN 0021-9258. PMID 15894804. doi:10.1074/jbc.M412494200. Процитовано 7 грудня 2020.
- Розділ 8.13. АНТИНОЦИЦЕПТИВНА СИСТЕМА.
- Goldstein, A.; Ghazarossian, V. E. (1980-10). Immunoreactive dynorphin in pituitary and brain. Proceedings of the National Academy of Sciences of the United States of America 77 (10). с. 6207–6210. ISSN 0027-8424. PMID 6108564. doi:10.1073/pnas.77.10.6207. Процитовано 7 грудня 2020.
- Drake, Carrie T.; Chavkin, Charles; Milner, Teresa A. (2007). Opioid systems in the dentate gyrus. Progress in Brain Research 163. с. 245–263. ISSN 0079-6123. PMID 17765723. doi:10.1016/S0079-6123(07)63015-5. Процитовано 7 грудня 2020.
- Goldstein, A.; Tachibana, S.; Lowney, L. I.; Hunkapiller, M.; Hood, L. (1979-12). Dynorphin-(1-13), an extraordinarily potent opioid peptide. Proceedings of the National Academy of Sciences of the United States of America 76 (12). с. 6666–6670. ISSN 0027-8424. PMID 230519. doi:10.1073/pnas.76.12.6666. Процитовано 7 грудня 2020.
- Lai, J.; Luo, Miaw-chyi; Chen, Q.; Ma, S.; Gardell, L.; Ossipov, M.; Porreca, F. (2006). Dynorphin A activates bradykinin receptors to maintain neuropathic pain. Nature Neuroscience. doi:10.1038/nn1804. Процитовано 7 грудня 2020.
- Merg, Florence; Filliol, D.; Usynin, I.; Bazov, I.; Bark, N.; Hurd, Y.; Yakovleva, T.; Kieffer, B. та ін. (2006). Big dynorphin as a putative endogenous ligand for the κ‐opioid receptor. Journal of neurochemistry. doi:10.1111/j.1471-4159.2006.03732.x. Процитовано 7 грудня 2020.
- Han, J. S.; Xie, C. W. (1984-02). Dynorphin: potent analgesic effect in spinal cord of the rat. Scientia Sinica. Series B, Chemical, Biological, Agricultural, Medical & Earth Sciences 27 (2). с. 169–177. ISSN 0253-5823. PMID 6147015. Процитовано 7 грудня 2020.
- Ren, M.; Lu, C.; Han, J. S. (1985). Dynorphin-A-(1–13) antagonizes morphine analgesia in the brain and potentiates morphine analgesia in the spinal cord. undefined (англ.). Процитовано 7 грудня 2020.
- J. Lai; Miaw-chyi Luo; Q. Chen; S. Ma; L. Gardell; M. Ossipov; F. Porreca (2006). Dynorphin A activates bradykinin receptors to maintain neuropathic pain. undefined (амер.). Процитовано 7 грудня 2020.
- Svensson, Camilla I.; Hua, Xiao-Ying; Powell, Henry C.; Lai, Josephine; Porreca, Frank; Yaksh, Tony L. (2005-10). Prostaglandin E2 release evoked by intrathecal dynorphin is dependent on spinal p38 mitogen activated protein kinase. Neuropeptides 39 (5). с. 485–494. ISSN 0143-4179. PMID 16176831. doi:10.1016/j.npep.2005.08.002. Процитовано 7 грудня 2020.
- Svensson, C.; Hua, X.; Protter, A.; Powell, H.; Yaksh, T. (2003). Spinal p38 MAP kinase is necessary for NMDA-induced spinal PGE2 release and thermal hyperalgesia. undefined (англ.). Процитовано 7 грудня 2020.
- Xu, Mei; Petraschka, Michael; McLaughlin, Jay P.; Westenbroek, Ruth E.; Caron, Marc G.; Lefkowitz, Robert J.; Czyzyk, Traci A.; Pintar, John E. та ін. (12 травня 2004). Neuropathic pain activates the endogenous kappa opioid system in mouse spinal cord and induces opioid receptor tolerance. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience 24 (19). с. 4576–4584. ISSN 1529-2401. PMC 2376823. PMID 15140929. doi:10.1523/JNEUROSCI.5552-03.2004. Процитовано 7 грудня 2020.
- Xu, Mei; Bruchas, Michael R.; Ippolito, Danielle L.; Gendron, Louis; Chavkin, Charles (7 березня 2007). Sciatic nerve ligation-induced proliferation of spinal cord astrocytes is mediated by kappa opioid activation of p38 mitogen-activated protein kinase. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience 27 (10). с. 2570–2581. ISSN 1529-2401. PMC 2104780. PMID 17344394. doi:10.1523/JNEUROSCI.3728-06.2007. Процитовано 7 грудня 2020.
- Sivam, S. P. (1989-09). Cocaine selectively increases striatonigral dynorphin levels by a dopaminergic mechanism. The Journal of Pharmacology and Experimental Therapeutics 250 (3). с. 818–824. ISSN 0022-3565. PMID 2476548. Процитовано 7 грудня 2020.
- Krebs, M. O.; Gauchy, C.; Desban, M.; Glowinski, J.; Kemel, M. L. (1994-04). Role of dynorphin and GABA in the inhibitory regulation of NMDA-induced dopamine release in striosome- and matrix-enriched areas of the rat striatum. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience 14 (4). с. 2435–2443. ISSN 0270-6474. PMC 6577130. PMID 7908960. Процитовано 7 грудня 2020.
- The 3rd meeting on the Therapeutic Potential of Kappa Opioids in Pain and Addiction was held at the Carolina Inn in Chapel Hill, NC.
- Cahill, Catherine M.; Taylor, Anna M. W.; Cook, Christopher; Ong, Edmund; Morón, Jose A.; Evans, Christopher J. (2014). Does the kappa opioid receptor system contribute to pain aversion?. Frontiers in Pharmacology (English) 5. ISSN 1663-9812. PMC PMC4233910. PMID 25452729. doi:10.3389/fphar.2014.00253. Процитовано 7 грудня 2020.
- Brugos, Brugos; Arya, Vikram; Hochhaus, Guenther (28 грудня 2004). Stabilized dynorphin derivatives for modulating antinociceptive activity in morphine tolerant rats: effect of different routes of administration. The AAPS journal 6 (4). с. e36. ISSN 1550-7416. PMC 2751232. PMID 15760101. doi:10.1208/aapsj060436. Процитовано 7 грудня 2020.