Махові пера
Махові пера — довгі жорсткі симетрично розташовані пера крил птахів. Головною функцією цих пір'їн є утворення підйомної сили та тяги, зумовлюючи таким чином здатність птахів до польоту. Махові пера деяких птахів також виконують й інші функції, зазвичай пов'язані із статевою поведінкою або живленням. Так, у деяких видів ці пера розвинулися у довге кольорове пір'я, а у інших пір'їни видають звуки в польоті. Маленькі насічки на передньому краю пір'я допомагають совам літати дуже тихо та в результаті успішно полювати. Навіть нелітаючі птахи мають махові пера, хоча часто у дуже видозміненій формі.
.jpg.webp)
Линяння махових пір'їн часто викликає серйозні проблеми для птахів, обмежуючи їх здатність до польоту. Різні види розвинули різні стратегії запобігання незахищеності протягом цього періоду, від скидання всього пір'я одночасно, зменшуючи період нездатності до польоту, до тривалого періоду линяння, що може займати до кількох років.
Махові пера крил
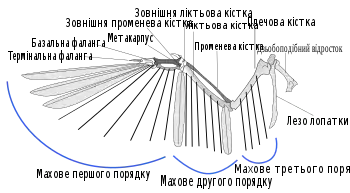
Махові пера крил (лат. remiges) розташовані на задньому (постеріорному) краю крил. Зв'язки прикріпляють його до кісток, а товстий шар сухожилля забезпечує їх жорстке положення[1]. Махові пера птахів симетричні на обох крилах, тотожні за структурою і формою (окрім випадків мутацій та травм), але не обов'язково за забарвленням[2][3]. Залежно від розташування, це пір'я поділяється на кілька груп, розташованих уздовж крила.
Першорядні махові
.jpg.webp)
Першорядні махові пера прикріпляються до кісток мануса («кисті» птахів, що складається з карометакарпуса і фаланги); це найдовше та найвужче пір'я крил птаха (особливо те, що прикріплене до фаланги), причому кожне перо може незалежно обертатися. Це пір'я особливо важливе для махового польоту та є головним джерелом тяги, воно штовхає птаха вперед. Тяга переважно утворюється при русі униз під час махового польоту. При русі уверх (коли крило рухається ближче до тіла птаха) махове першого порядку розходиться та обертається для зменшення аеродинамічного опору, проте надаючи додаткову тягу[4]. Гнучкість пір'я крил також дозволяє великим птахам ширяти з розставленими маховими першого порядку для зменшення завихрень на кінці крила і зменшення аеродинамічного опору[5].
Різні види птахів істотно розрізняються за числом махових першого порядку. Так, більшість негоробцеподібних мають 9 або 11 на кожному крилі[6], хоча пірникозові, лелекоподібні і фламінго мають 12[7], а страуси мають 16[7]. Більшість сучасних горобцеподібних мають 10 махових першого порядку[6], а деякі 9. Ті, що мають 9, бракують найбільш дистального пера, зазвичай коротшого і часто рудиментарного у горобцеподібних[7].
Другорядні махові пера

Махові пера другого порядку приєднано до ліктьової кістки. У деяких видів птахів зв'язки, що сполучають махове крил, приєднані до невеликих проекцій цієї кістки, відомих як пір'єві вузли; у інших видів з'єдання пряме, а цих вузлів не існує. Махові другого порядку птах тримає разом протягом польоту, це пір'я нездатне то розведення, як махові першого порядку, та допомагає птаху утворювати підіймальну силу через надання аеродинамічної форми крилу. Махові другого порядку зазвичай коротші та ширші за махові першого порядку, із тупішими кінцями. Птахи мають від 6 махових другого порядку у колібрі до 40 у деяких видів альбатросів[8]. Зазвичай більші види з довшими крилами мають більше число цих пір'їн[8].
Птахи з понад 40 родин негоробцеподібних птахів не мають п'ятого пера на кожному крилі, явище, відоме як діастраксія[9]. До цієї групи належать гагари, пірникози, пелікани, яструби, орли, журавлі, баранці, мартини, папуги і сови. У цих птахів п'ятий набір криючого пір'я другого порядку не вкриває махове, ймовірно через поворот пір'я протягом ембріонального розвитку[9].
Третьорядні махові пера
Махові пера третього порядку прикріплені до плечової кістки та наявні лише у деяких видів. Багато дослідників загалом не використовують термін «махове третього порядку», розглядаючи це пір'я як частину махового другого порядку. Це пір'я діє як захисний покрив для згорнутих махових першого та другого порядку[10].
Виїмки
Найбільш дистальні (найдальші) махові першого порядку великих птахів, перш за все хижих птахів, часто мають чітко виражені виїмки у різних частинах краю пера. Ці виїмки досить сильно розрізняються за розташуванням та глибиною[9]. Тоді як неглибокі виїмки можуть знаходитися на будь-якій стороні пера, найглибші завжди знаходяться на ширшій задній стороні пера. Ці виїмки можна побачити на зображенні пір'їн приблизно посередині обох боків пера, що зображене ліворуч — глибокі на лівій стороні та неглибокі на правій. Функція виїмок все ще залишається спірною. Для її пояснення було запропоновано кілька гіпотез, які, проте, є лише теоретичними, не підкріпленими достатніми експериментальними даними[11]. Зокрема, було запропоновано гіпотезу, що наявність цих виїмок створює щілини на краю крила, що покращує аеродинамічні властивості крила, зокрема збільшуючи підіймальну силу, зменшуючи силу аеродинамічного опору[12] та збільшуючи стабільність у польоті[13].
Придаткове крило

Пір'я придаткового крила зазвичай не вважається маховим — хоча це пір'я асиметричне, воно не має жорсткості, характерної для махового. Проте, пір'я придаткового крила відіграє важливу роль у польоті, допомагаючи при повільному русі. Це пір'я прикріплене до кісток великого пальця птаха і зазвичай розташоване на передній стороні крила подібно до передкрилків крила літаків, дозволяючи крилу досягати більшого кута атаки та збільшення підіймаючої сили, запобігаючи втраті швидкості. Рухаючи великий палець та змінюючи щілину між придатковим крилом та рештою крила, птах запобігає втраті швидкості, що необхідно при повільному польоті або приземленні[9].
Затримка розвитку в гоацинів
Розвиток махових пер крил та придаткового крила у гоацинів значно уповільнений порівняно з рештою молодих птахів, ймовірно через наявність у молодих гоацинів кігтів на перших двох пальцях. Ці кігті використовуються, щоб лазити по деревах, а розвиток пір'я заважає цій функції. Зазвичай ці птахи відкидають кігті між 70-м і 100-м днями життя, але у деяких вони залишаються і в дорослому стані[14][15].
Посилання
- Podulka, Sandy, Ronald W. Rohrbaugh and Rick Bonney, ред. (2003). Home Study Course in Bird Biology, Second Edition. Ithaca, NY: Cornell Lab of Ornithology. с. 1.11.
- Trail, 2001, p. 8
- Moller, Anders Pape; Jacob Hoglund (1991). Patterns of Fluctuating Asymmetry in Avian Feather Ornaments: Implications for Models of Sexual Selection. Proceedings: Biological Sciences 245 (1312): 1–5. doi:10.1098/rspb.1991.0080.
- Ehrlich et al., 1994, p. 219
- Ehrlich et al., 1994, p. 79
- Jenni, Lukas; Winkler, Raffael (1994). Moult and Ageing of European Passerines. London: Academic Press. ISBN 0-12-384150-X.
- del Hoyo, Elliott & Sargatal, 1992, p. 37
- Sibley, 2001, p. 17
- Campbell, Bruce; Lack, Elizabeth, eds. (1985). A Dictionary of Birds. Carlton, England: T and A D Poyser. ISBN 0-85661-039-9., page 656
- Ferguson-Lees & Christie, 2001, p. 27
- John Videler (2005). Avian Flight. Oxford University Press. ISBN 978-0-19856603-8.
- Trail, 2001, p. 6
- Norberg (1990). Verterbrate Flight. Berlin: Springer.
- Campbell & Lack, 1985, p. 285
- del Hoyo, Elliott & Sargatal, 1996, p. 29
Література
- Trail, Pepper (2001). Wing Feathers (PDF). U.S. Fish and Wildlife Service. Архів оригіналу за 15 липня 2008. Процитовано 30 травня 2008.
- Ehrlich, Paul R.; Dobkin, Darryl A.; Wheye, Darryl; Pimm, Stuart L. (1994). The Birdwatcher's Handbook. Oxford University Press. ISBN 0-19-858407-5.
- Ferguson-Lees, James; Christie, David A. (2001). Raptors of the World. London: Christopher Helm. ISBN 0-7136-8026-1.
- Campbell, Bruce; Lack, Elizabeth, ред. (1985). A Dictionary of Birds. Carlton, England: T and A D Poyser. ISBN 0-85661-039-9.
- del Hoyo Josep, Elliott Andrew, Sargatal Jordi, ред. (1992). Handbook of the Birds of the World, Volume 1: Ostrich to Ducks. Barcelona: Lynx Edicions. ISBN 84-87334-10-5.