М'язове скорочення
М'язове скорочення — реакція м'язових клітин на вплив нейромедіатора, рідше гормону, що виявляється в зменшенні довжини клітини. Це життєво важлива функція організму, пов'язана з оборонними, дихальними, харчовими, статевими, видільними і іншими фізіологічними процесами.
Всі види довільних рухів — ходьба, міміка, руху очних яблук, ковтання, дихання і т. ін. здійснюються за рахунок скелетних м'язів . Мимовільні рухи (крім скорочення серця) — перистальтика шлунка і кишечника, зміна тонусу кровоносних судин, підтримання тонусу сечового міхура — обумовлені скороченням гладкої мускулатури . Робота серця забезпечується скороченням серцевої мускулатури .
Типи
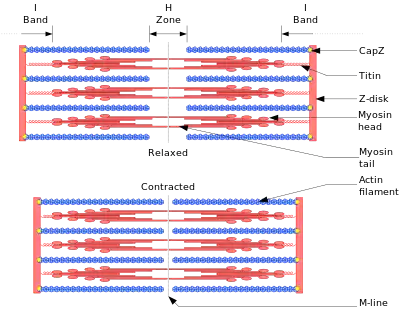
Основою всіх типів м'язового скорочення служить взаємодія актину і міозину . У скелетних м'язах за скорочення відповідають міофібрили (приблизно дві третини сухої ваги м'язів). Міофібрили — структури товщиною 1-2 мкм, що складаються з саркомерів — структур довжиною близько 2,5 мкм, що складаються з Актинові і міозінових (тонких і товстих) філаментів і Z-дисків, з'єднаних з Актинові філаменти. Скорочення відбувається при збільшенні концентрації в цитоплазмі іонів Ca 2+ в результаті ковзання міозінових філаментів щодо Актинові. Джерелом енергії скорочення служить АТФ. ККД м'язової клітини близько 50 %, м'язи в цілому не більше 20 %. Максимальна сила м'язів не досягається в реальних умовах; не всі клітини м'язи використовуються одночасно і скорочуються з максимальною силою, інакше, при скороченні багатьох скелетних м'язів будуть пошкоджені сухожилля або кістки (що іноді і спостерігається при сильних судомах). ККД м'язи також залежить від зовнішніх умов; наприклад, на холоді він значно знижується, так як для організму важливіше зберегти температуру тіла[1] .
Ізометричне скорочення
Ізометричне скорочення м'язи створює напругу без зміни довжини. Приклад можна знайти, коли м'язи руки і передпліччя захоплюють об'єкт; суглоби руки не рухаються, але м'язи генерують достатню силу, щоб запобігти падінню об'єкта.
Ізотонічне скорочення
При фізіологічному скороченні напруга в м'язах залишається постійною, незважаючи на зміну довжини м'язів. Це відбувається, коли сила скорочення м'язів відповідає загальному навантаженню на м'язи.
Концентричне скорочення
При концентричному скороченні м'язової напруги достатньо для подолання навантаження, а м'яз коротшає при скороченні. Це відбувається, коли сила, створювана м'язом, перевищує навантаження, протидіє її скороченню.
Ексцентричне скорочення
При ексцентричному скороченні напруга є недостатньою для подолання зовнішнього навантаження на м'язи, і м'язові волокна подовжуються при їх скороченні. Замість того, щоб тягнути суглоб в напрямку скорочення м'язів, м'яз діє так, щоб уповільнити суглоб в кінці руху або іншим чином контролювати переміщення вантажу. Це може відбуватися мимоволі (наприклад, при спробі перемістити вагу, занадто важкий для підйому м'язи) або добровільно (наприклад, коли м'яз «згладжує» рух або чинить опір гравітації, як, наприклад, під час ходьби вниз). У короткостроковій перспективі силові тренування за участю як ексцентричних, так і концентричних скорочень, мабуть, збільшують м'язову силу більше, ніж тренування тільки з концентричними скороченнями. Однак викликане фізичними вправами пошкодження м'язів також більше при подовженні скорочень.
Ексцентричні скорочення в русі
Ексцентричні скорочення зазвичай виникають як гальмуючі зусилля на противагу концентричному стиску, щоб захистити суглоби від пошкодження. Під час практично будь-якого звичайного руху ексцентричні скорочення допомагають зберігати плавність рухів, але також можуть уповільнювати швидкі рухи, такі як удар або кидок. Частина тренування для швидких рухів, таких як качки під час бейсболу, включає в себе зменшення ексцентричного гальмування, що дозволяє розвивати велику потужність під час руху.
Ковзання міозину відносно актину
Головки міозину розщеплюють АТФ і за рахунок енергії, що вивільняється, змінюють конформацію, ковзаючи по Актинові филаментам. Цикл можна розділити на 4 стадії:
- Вільна головка міозину зв'язується з АТФ і гідролізує його до АДФ і фосфату і залишається пов'язаної з ними. (Оборотний процес — енергія, що виділилася в результаті гідролізу, запасається в зміненій конформації міозину).
- Головки слабо зв'язуються з наступною субодиницею актину, фосфат відокремлюється, і це призводить до міцного зв'язування головки міозину з Актинові філаментів. Ця реакція вже необоротна.
- Головка зазнає конформационное зміна, яка провадить підтягування товстого філамента до Z-диску (або, що еквівалентно, вільних кінців тонких філаментів один до одного).
- Відділяється АДФ, за рахунок цього головка відділяється від актинового філамента. Приєднується нова молекула АТФ.
Далі цикл повторюється до зменшення концентрації іонів Ca 2+ або вичерпання запасу АТФ (в результаті смерті клітини). Швидкість ковзання міозину по актину ≈ 15 мкм/с. У міозіновому філаменті багато (близько 500) молекул міозину і, отже, при скороченні цикл повторюється сотнями головок відразу, що і призводить до швидкого і сильного скорочення. Слід зауважити, що міозин поводиться як фермент — актин-залежна АТФаза. Так як кожне повторення циклу пов'язано з гідролізом АТФ, а отже, з позитивною зміною вільної енергії, то процес односпрямований. Міозин рухається по актину тільки в сторону плюс-кінця.
Для скорочення м'яза використовується енергія гідролізу АТФ, але м'язова клітина має вкрай ефективну систему регенерації запасу АТФ, так що в розслабленому і працюючому м'язі зміст АТФ приблизно рівний. Фермент фосфокреатінкіназа каталізує реакцію між АДФ і креатинфосфату, продукти якої — АТФ і креатин . Креатинфосфат містить більше запасеної енергії, ніж АТФ. Завдяки цьому механізму при спалаху активності в м'язовій клітці падає зміст саме креатинфосфату, а кількість універсального джерела енергії — АТФ — не змінюється. Механізми регенерації запасу АТФ можуть відрізнятися в залежності від парціального тиску кисню в оточуючих тканинах (див. Анаероби).
Механізм регуляції
В основному в регуляції м'язової активності беруть участь нейрони, але є випадки, коли скороченням гладкої мускулатури управляють і гормони (наприклад, адреналін і окситоцин). Сигнал про скорочення можна розділити на кілька етапів:
Від клітинної мембрани до саркоплазматичного ретикулума
Вплив медіатора, що виділився з мотонейрона, викликає потенціал дії на клітинній мембрані м'язової клітки, який передається далі за допомогою спеціальних впячіваній мембрани, званих трубочками, які відходять від мембрани всередину клітини. Від Т-трубочок сигнал передається саркоплазматичний- особливому компартмент з сплощені мембранних бульбашок (ендоплазматичноїмережі м'язової клітини), що оточують кожну міофібрил. Цей сигнал викликає відкриття Ca 2+ -каналів в мембрані ретикулума. Назад іони Ca 2+ потрапляють в ретикулум за допомогою мембранних кальцієвих насосів — Ca 2+ — АТФази .
Від виділення іонів Ca 2+ до скорочення міофібрил
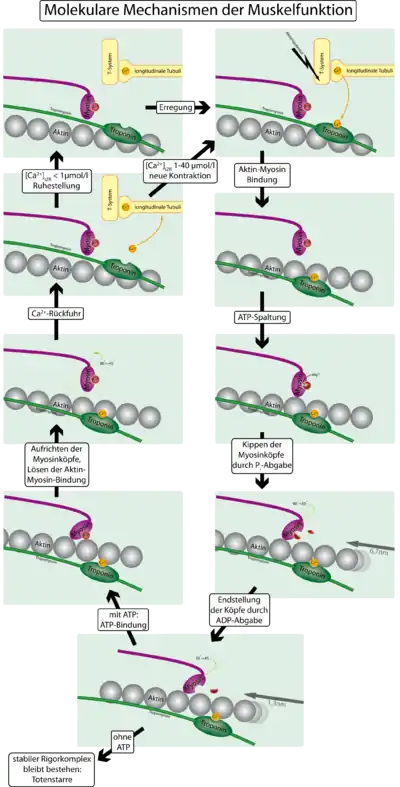
Для того, щоб контролювати скорочення, до Актинові філаменти прикріплюється білок тропомиозин і комплекс з трьох білків — тропонин (субодиниці цього комплексу називаються тропоніну T, I і C). Тропонин C — близький гомолог іншого білка, кальмодулина . Через кожні сім субодиниць актину розташований тільки один тропоніновим комплекс. Зв'язок актину з тропонином I переміщує тропомиозин в положення, що заважає зв'язку міозину з актином. Тропонин C зв'язується з чотирма іонами Ca 2+ і послаблює дію тропоніну I на актин, і тропомиозин займає положення, яке не перешкоджає зв'язку актину з міозином. Джерелом енергії для скорочення м'язових волокон служить АТФ. При зв'язуванні тропонина з іонами кальцію активуються каталітичні центри для розщеплення АТФ на голівках міозину. За рахунок ферментативної активності головок міозину гідролізується АТФ, розташований на голівці міозину, що забезпечує енергією зміна конформації головок і ковзання ниток. Звільнитися при гідролізі АТФ молекула АДФ і неорганічний фосфат використовуються для подальшого ресинтезу АТФ. До міозіновой голівці приєднується нова молекула АТФ. При цьому відбувається роз'єднання поперечного містка з ниткою актину. Повторне прикріплення і від'єднання містків триває до тих пір, поки концентрація кальцію всередині міофібрил не знизиться до подпороговой величини. Тоді м'язові волокна починають розслаблятися.
Основні білки міофібрил
| білок | Частка білка% | Його мовляв. маса, kDa | його функція |
|---|---|---|---|
| міозин | Головний компонент товстих філаментів. Утворює зв'язку з актином. Рухається по актину за рахунок гідролізу АТФ. | ||
| актин | Головний компонент тонких філаментів. Під час скорочення м'язи по ньому рухається міозин. | ||
| Тітин | Великий гнучкий білок, який утворює ланцюг для зв'язування міозину з Z-диском. | ||
| Тропонін | Комплекс з трьох білків, що регулює скорочення при зв'язуванні з іонами Ca 2+. | ||
| Тропоміозин | Пов'язаний з Актинові філаменти стрижневидний білок, який блокує рух міозину. | ||
| Небулін [en] | Довгий нерозтяжної білок, пов'язаний з Z-диском і йде паралельно Актинові филаментам. |
Примітки
Література
- Б. Альбертс, Д. Брей, Дж. Льюїс, М. Рефф, К. Робертс, Дж. Уотсон, Молекулярна біологія клітини — В 3-х т. — Пер. з англ. — Т.2. — М .: Світ, 1994. — 540 с.
- М. Б. Беркінбліт, С. М. Глаголєв, В. А. Фуралев, Загальна біологія — У 2-х ч. — Ч.1. — М.: МИРОС, 1999. — 224 с .: іл.
Посилання
- Скорочення м'яза // Універсальний словник-енциклопедія. — 4-те вид. — К. : Тека, 2006.
