Ендоплазматичний ретикулум
Ендоплазматичний ретикулум (ЕПР, від лат. reticulum — «сіточка») або ендоплазматична мережа — внутрішньоклітинна органела еукаріотичних клітин, що являє собою розгалужену систему з оточених мембраною сплющених порожнин, бульбашок і канальців.
Історія відкриття
Вперше ендоплазматичний ретикулум був виявлений канадськими вченими Кітом Портером, Альбертом Клодом і Ернстом Фулламом в 1945 році за допомогою електронної мікроскопії[1].
Будова
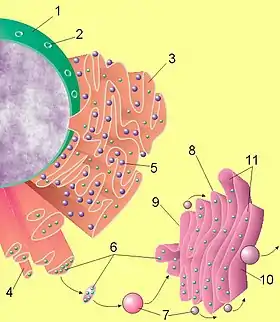
Ендоплазматичний ретикулум складається з розгалуженої мережі трубочок і кишень, оточених мембраною. Площа мембран ендоплазматичного ретикулума складає більше половини загальної площі всіх мембран клітини.
Мембрана ЕПР морфологічно ідентична оболонці клітинного ядра і складає з нею одне ціле. Таким чином, порожнини ендоплазматичного ретикулума відкриваються в міжмембранну порожнину ядерної оболонки. Мембрани ЕПС забезпечують активний транспорт ряду елементів проти градієнту концентрації. Бульбашки та канальці, що створюють ендоплазматичний ретикулум, мають в поперечнику 0,05-0,1 мікрона (іноді до 0,3 мікрона), товщина двошарових мембран, що стінку канальців, становить близько 50 ангстрем. Ці структури містять ненасичені фосфоліпіди, а також деяку кількість холестерину і сфінголіпідів. У їхній склад також входять білки. Найтонші трубочки, діаметр яких коливається в межах 1000-3000 ангстрем, заповнені гомогенним вмістом та сполучають більші за розміром частини ендоплазматичного ретикулума. Подібно до компонентів цитоскелета мембрани полярні: з одного кінця вони нарощуються, а з іншого- розпадаються на окремі фрагменти. Ендоплазматичний ретикулум не є стабільною структурою і схильний до частих змін. Виділяють два типи ЕПР:
- Шорсткий (гранулярний) ендоплазматичний ретикулум,
- Гладкий (агранулярний) ендоплазматичний ретикулум.
На поверхні шорсткого ендоплазматичного ретикулума знаходиться велика кількість рибосом, які відсутні на поверхні гладкого ЕПР.
Шорсткий та гладкий ендоплазматичний ретикулум виконують деякі різні функції в клітині.
Функції ендоплазматичного ретикулума
За участю ендоплазматичного ретикулума відбувається трансляція та транспорт мембранних білків, що секретуються, синтез і транспорт ліпідів і стероїдів. Для ЕПС характерно також накопичення продуктів синтезу. Ендоплазматичний ретикулум бере участь у тому числі і в створенні нової ядерної оболонки (наприклад після мітозу). Ендоплазматичний ретикулум містить внутріклітинний запас кальцію, який є медіатором багатьох реакцій відповіді клітини, зокрема скорочення м'язових клітин. У клітинах м'язових волокон розташована особлива форма ендоплазматичного ретикулума — саркоплазматичний ретикулум.
Функції гладкого ендоплазматичного ретикулума
Гладкий ендоплазматичний ретикулум бере участь в багатьох процесах метаболізму. Ферменти гладкого ендоплазматичного ретикулума беруть участь у синтезі ліпідів і фосфоліпідів, жирних кислот і стероїдів. Також агранулярний ендоплазматичний ретикулум грає важливу роль у вуглеводному обміні, знезараженні клітки і запасанні кальцію. Зокрема, у зв'язку з цим в клітках надниркових залоз і печінки переважає гладкий ендоплазматичний ретикулум.
Синтез гормонів
До гормонів, які утворюються в гладкому ЕПС, належать, наприклад, статеві гормони хребетних тварин і стероїдні гормони (що синтезуються в надниркових залозах). Клітини яєчок і яєчників, що відповідають за синтез гормонів, містять велику кількість гладкого ЕПР.
Накопичення і перетворення вуглеводів
Вуглеводи в організмі накопичуються в печінці у вигляді глікогену. За допомогою гліколізу глікоген в печінці трансформується в глюкозу, що є найважливішим процесом в підтримці рівня глюкози в крові. Один з ферментів гладкого ЕПС відщеплює від першого продукту гліколізу, глюкоза-6-фосфату, фосфогрупу, дозволяючи, таким чином, глюкозі залишити клітину і підвищити рівень цукру в крові.
Нейтралізація отрут
Гладенький ендоплазматичний ретикулум клітин печінки бере активну участь в нейтралізації токсинів. Ферменти гладенького ЕПР приєднують до нерозчинних радикалів гідрофільні послідовності, внаслідок чого токсин стає краще розчинним і може виводитися з клітини, а згодом і з організму через сечу. У разі безперервного надходження отрут, медикаментів або алкоголю, площа гладенького ЕПР збільшується, що дозволяє нейтралізувати більшу кількість токсину за рахунок збільшення робочої поверхні ЕПР.
Саркоплазматичний ретикулум
Особлива форма гладкого ендоплазматичного ретикулума, саркоплазматичний ретикулум, утворюється в м'язових клітинах, де іони кальцію активно закачуються з цитоплазми в порожнині ЕПР проти градієнту концентрації в не збудженому стані клітини і звільняються в цитоплазму для ініціації скорочення. Концентрація іонів кальцію в ЕПС може досягати 10−3 моль/л, тоді як в цитоплазмі — порядка 10−7 моль/л (в стані спокою). Таким чином, мембрана саркоплазматичного ретикулума забезпечує активне перенесення проти градієнтів концентрації великих порядків. І прийом і звільнення іонів кальцію в ЕПС знаходиться в жорсткій залежності від фізіологічних умов. 2
Концентрація іонів кальцію в цитозолі впливає на безліч внутрішньоклітинних і міжклітинних процесів, таких як активація або інгібування ферментів, експресія генів, синаптична пластичність нейронів, скорочення м'язових клітин, звільнення антитіл з клітин імунной системи.
Функції гранулярного ендоплазматичного ретикулума
Гранулярий ендоплазматичний ретикулум має дві функції: синтез білків і виробництво мембран.
Синтез білків
Білки, що виробляються клітиною, синтезуються в рибосомах, які можуть бути приєднані до поверхні ЕПС, саме це і надає поверхні «шорсткий», гранулярний вигляд[2]. На поверхні ЕПР синтезується багато білків, призначених для сортування і транслокації до певних відділів клітини, тобто інтегральні мембранні білки, білки органел і білки, призначені для секреції. Проте, рибосоми на поверхні ЕПР на завжди знаходяться там, а приєднуються до нього після початку синтезу білка, призначеного для сортування в ЕПР[3]. Вільна рибосома синтезує білок, поки цитозольні частинки розпізнавання сигналу на впізнають сигнал з 5-15 гідрофобних амінокислот (N-сигнальна послідовність) після позитивно зярядженої амінокислоти на початку ланцюжка. Цей сигнальний фрагмент залишається в мембрані до кінця синтезу, після чого він відрізається від готового білка.
Отримані поліпептідні ланцюги поміщаються в порожнини гранулярного ендоплазматичного ретикулума, де згодом піддаються першим крокам посттрансляційної модифікації і згортаються. Таким чином, лінійні ланцюги амінокислот отримують після транслокації в ендоплазматичний ретикулум необхідну тривимірну структуру.
Синтез мембран
За допомогою виробництва фосфоліпідів, ЕПР розширює поверхню своїх власних мембран, які за допомогою транспортних везикул транспортуються до інших частин мембранної системи.
Посилання
- Porter KR, Claude A, Fullam EF (1945 Mar). A study of tissue culture cells by electron microscopy. J Exp Med. 81: 233–246.
- Campbell, Neil A. (1996) Biology Fourth Edition. Benjamin/Cummings Publishing, pp. 120–121 ISBN 0-8053-1940-9
- Lodish, Harvey, et al. (2003) Molecular Cell Biology 5th Edition. W. H. Freeman, pp. 659–666 ISBN 0-7167-4366-3