Теорія острівної біогеографії
Моделі острівної біогеографії (англ. island biogeography) були запропоновані Робертом Макартуром і Е. О. Вілсоном (1967)[1] для пояснення того факту, що на кожному з K островів, що входять до складу архіпелага, мешкає менше видів, ніж на тій же території, що входить до складу материка, з тими ж місцепроживаннями. S1, S2, S3 … Sk <S0 числа видів на тій же території материка і менше P, де P — загальне число видів даної групи, що мешкають на даній території, видовий фонд, з якого рекрутуються види для заселення островів і/або перерозподілу всередині материка.
Основні положення
Зменшення Sk в порівнянні з S0 було названо острівним ефектом: він визначається як ймовірність випадання на островах архіпелагу якоїсь частини видів вихідної фауни або флори, які постійно мешкають на материку. Також важливо, що схильність до острівного ефекту і життя на архіпелазі інших (тих, що не випали) видів означає сугубу нестійкість існування — і протиставляється стійкості їх же на материку. Збережені види весь час «стрибають з крижини на крижину» — їх поселення вимирає на одних островах і з'являються на інших в процесі реколонізації, так що населення виду активно перерозподіляється в острівному ареалі. Тому модель МакАртура-Вілсона називається рівноважною.
Природно, це підвищує ймовірність вимирання виду на архіпелазі в цілому, якщо якийсь «стрибок» раптом виявиться невдалим, особливо якщо довгий час переміщень не відбувається, і поселення виду концентруються на якомусь — скажімо, наприклад, найбільшому — з островів. Тут вони найвразливіші.
У вихідній моделі Мак-Артура — Вілсона ступінь ізоляції острову від джерела нових видів — більшого острова або материка — впливала тільки на темп імміграції і, відповідно, на швидкість колонізації острова видами, які раніше випали зі складу фауни (або зовсім новими). Те, що ізольованість островів настільки ж, якщо не більше, сильно, впливає на ризик вимирання видових поселень, «замкнених» на островах — чим більше ізольованість, тим вище ризик, причому залежність ступенева — це доповнення кінця 1980-х рр.
Також моделі розглядають процеси екологічних, а не «еволюційних масштабів часу, тобто не стосуються адаптивної радіації нових видів від предківської форми, що розселилися по всіх островах, як у випадку з дарвіновими в'юрками. Максимум, про що можна говорити, це перші стадії мікроеволюції; взагалі, зараз вже абсолютно зрозуміло, що екологічні й еволюційні процеси йдуть одночасно і або посилюють, або послаблюють один одного. Пристосування осіб, скажімо, до повернення старого хижака, забруднення водойми, пожеж та інших середовищних стресів супроводжується елементарними еволюційними явищами в популяції, що об'єднує їх.
Скажімо, пристосувальні трансформації популяційної системи виду, так щоб остання могла стійко відтворюватися на островах, а не тільки на материку і, таким чином, робилася нечутливою до острівного ефекту. Про ці процеси (названі зворотною урбанізацією) мова піде далі, вони найбільш значимі при заселенні „архіпелагів“ лісових, лучних, водно-болотних і тощо природних середовищ існування, що виникли під дією антропогенної фрагментації „дикими“ видами різних груп біоти, і в першу чергу тими, що були витіснені з „архіпелагу“ в процесі його утворення. Ці штучні „острови“ за ступенем ізольованості не поступаються природним, хоча найчастіше знаходяться на меншій дистанції, а їх місцеперебування зазвичай сильно антропогенно-трансформовані з країв (крайовий ефект, edge effect).
Острівний ефект зафіксовано для всіх груп біоти — птахів, метеликів, ссавців, безхребетних і павуків, амфібій, рептилій, риб в озерах, квіткових рослин та ін. У серії ретельних досліджень було показано, що зменшення числа видів на островах архіпелагу Sk в порівнянні з материком пов'язане саме з роздроблення на острови і з ізоляцією окремих островів, відстанню і іншими бар'єрами, а не тільки з меншою різноманітністю місць існування на меншій площі. Тобто останній ефект теж присутній, і домішується до острівного, але його можна відокремити, і острівний залишиться у всій силі.
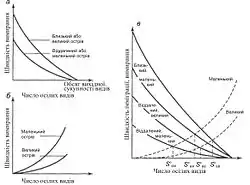
Згідно з вихідним, найпростішим варіантом моделі, число видів Sk на кожному з островів архіпелагу, визначається рівновагою між вимиранням і імміграцією, з материка, або більш великих островів. Сама рівновага має динамічний характер: види безперервно вимирають і заміщуються за рахунок реколонізації тими ж або новими видами.
Міркування, на яких ґрунтувалися МакАртур і Вілсон, були прості. „… На острові швидкість вимирання видів вища, ніж на материку, а швидкість заселення новими не настільки висока, щоб швидко компенсувати вимирання. Число видів, що входять одночасно в острівний біоценоз, є результатом збалансованості двох процесів, а саме — вселення (імміграції, I) нових видів на острів і вимирання (E) вже присутніх.
Обидва процеси можна виразити як функції числа видів, наявних на острові. Швидкість вселення знижується, оскільки все менше видів з вихідного пулу (що живуть на прилеглому материку чи великому острові) залишиться за межами острова. Швидкість же вимирання зростає — адже буде кому вимирати, а крім того, скорочення середнього розміру однієї популяції автоматично рівносильне підвищенню ймовірності її випадкового вимирання“.
Тому, хоча число видів на острові в цілому стабілізується на рівні Sk <S0; ця стабілізація — результат не сталості видового складу, а безперервної зміни видів, коли одні вимирають, а інші вселяються. Відповідно, на великих островах буде більше видів, ніж на дрібних (при тому, що на кожному — менше, ніж у середині материка), а видове багатство знижується в міру віддалення острова від джерела заселення. Тому форма архіпелагу і його орієнтація щодо материка також істотні: один результат буде у випадку, коли острів є продовженням материкового мису, півострова чи довгого виступу, який звужується, і інший — коли берегова лінія материка повернута до архіпелагу своєю довгою стороною.
Якщо острів лінзоподібний або витягнутий, для потрапляння видів на нього істотно, довгий чи короткий радіус паралельний узбережжю материка. Останнє ще вірогідніше для міських парків, лісових мікрофрагментів та інших „островів“, які утворилися внаслідок антропогенної фрагментації природних ландшафтів.
Теорія острівної біогеографії і антропогенна фрагментація ландшафтів
Скрізь вище йшлося про справжні острови, архіпелаги, материки і т. д. Однак зрозуміла (і виправдана) спокуса докласти моделі МакАртура-Вілсона до острівних місцепроживань, які утворилися внаслідок антропогенної фрагментації, якщо вона годиться для острівних місцеперебувань типу озер, печер, гірських лісів тощо. Дійсно, природоохоронці „кігтями і зубами“ вчепилися в неї і з початку 1980-х рр. І стали активно використовувати для прогнозу збереженості видів у заповідниках, заказниках, і інших особливо охоронюваних природних територіях (ООПТ).
Справа в тому, що починаючи з „ економічного буму“ 1950-1960-х рр. (максимум відносного приросту ВВП в ХХ столітті і момент появи нинішніх екологічних проблем) масиви хоч якось збережених природних ландшафтів в староосвоєних районах Європи, Східної Азії, Північної Америки і т. д. являють собою якраз „острови“, і досить ізольовані. Навіть малопорушені масиви тайгових лісів (які розвиваються в спонтанному режимі і зберегли специфічну ярусно-віконну мозаїку і пр.) являють собою „острови“, розділені масою вторинних лісів; і суцільного тайгового пояса більше не існує. Такий же „архіпелаг“ з „уламків“ природних ландшафтів різного ступеня зміненності виникає в процесі урбанізації регіона в силу двох головних закономірностей, які підтримують і підсилюють одна одну:
- У всіх досліджених агломераціях людність урбанізованого „ядра“ регіону зростає повільніше площі (Стольберг, 2001)[2]. Іншими словами, все більша частина міських функцій не може виконуватися на міській території. Вона все більше обслуговує зростаючий споживчий тиск городян — метраж квартир, зали магазинів та місць розваг, гаражі та вулиці для їхніх машин, і т. д., так що для звалищ, станцій водопідготовки, відстійників тощо загальнозначущих служб міста захоплюються все більші території регіону. Та й для будівництва житла теж.
- З 1970-80-х рр. у великих містах, а тим більше мегаполісах фіксується позитивний зв'язок між людністю міста і числом годин перебування середнього жителя на природі. Серед людей, які концентруюються в містах, підсилюється потяг до дикої природи і як компенсація, перебування там все тісніше зв'язується з „відпочинком“ і т. д.[3][4] Виникає і поширюється потік городян, які масово виїжджають на природу у відпустку і уїк-енд, що приносить їм користь — за підрахунками економістів, в середньому 20 днів на природі влітку ліквідують 3 дні непрацездатності з грипу взимку. Діючий при капіталізмі пріоритет приватного а/т над гроомадським, автомашин над ж/д робить цей потік виключно потужним агентом територіального зростання міст за рахунок субурбанізації, і дроблення природних ландшафтів периферії за рахунок рекреації (особливо якщо остання пов'язана з появою дач).
Опосередкованою ланкою в цій антропогенній фрагментації слугує прискорений розвиток мережі а/д, що ніби розносять вплив міста по регіону і одночасно фрагментують природні місцепроживання, а згодом „по тому ж руслу“ йдуть процеси урбанізації, вони змушують рекреантів від'їжджати ще далі і т.д . І дійсно, в цілому по країні руйнування природних ландшафтів тісно пов'язане з транспортною доступністю відповідних територій, і в першу чергу — шосейною. Відповідно, рекреаційне навантаження на регіон за наслідками у вигляді створення „архіпелагу“ (фрагментації великих масивів природних ландшафтів ззовні з інсулярізацією їх дорожньо-стежковою мережею зсередини) порівнюється з господарським освоєнням території.
Це дозволяє розширити „область визначення“ моделей острівної біогеографії, поширивши її з справжніх островів на „архіпелаги“, утворені „клаптиками“ природних ландшафтів (лісових, лучних, болотних і ін.), що збереглися при антропогенній фрагментації вихідного „материка“. Там дані елементи були „точно підігнані“ один до одного „в клаптикову ковдру“, і специфічний паттерн мозаїки „клаптиків“ показував непорушеність місцеперебування. Антропогенні порушення „розривають“ вихідний ландшафт, переважно винищують (або скорочують) окремі „клаптики“, і завжди ізолюють їх один від одного (разом з елементами вихідної мозаїки, які краще збереглися).
Так, для 25 дуже різних міст Європи, вивчених в рамках створення Європейського атласу урболандшафтів, було характерне територіальне розростання, з першочерговим витісненням з міста а) боліт, б) ріллі, в) відкритих трав'янистих місць існування. Лісові та лісопаркові ділянки витіснялися істотно повільніше (зберігалися найкраще на території к. НДР і ЧРСР — лідерів збереження біорізноманіття в місті в 1960-80-х рр.). Те ж вірно для природи Москви[5]
Тобто по мірі господарського освоєння території на місці вихідного „материка“ непорушених природних ландшафтів виникає „архіпелаг“ більш-менш збережених фрагментів останніх, завжди перетворених з країв і розділених матриксом з антропогенно-змінених територій, сільськогосподарських, гірничорудних і міських. Це дозволяє класифікувати різні типи „архіпелагів“, створених антропогенною фрагментацією середовища існування, розмістивши їх в двокоординатному просторі, одна вісь якого відображає швидкість процесу, а інша — ступінь роздробленості і трансформування фрагментів вихідних місць існування, що збереглися на даний момент. Важливі приклади антропогенних „архіпелагів“:
- Лісові мікрофрагменти в степовій і лісостеповій зоні, а також в „антропогенному лісостепу“;
- „Архіпелаг“ верхових боліт на Поліссі та інших промислово розвинених областях, де йшло інтенсивне осушення і торфорозробки, особливо фрезерним способом і методом гідроторфа;
- Лісові і лукові масиви, ізольовані один від одного мережею автодоріг з досить інтенсивним рухом а/т;
- „Острова“ сільгоспугідь, полів і лук, різною мірою підрозділені зсередини завдяки заболочуванню і заростанню кущами, ізольовані один від одного населеними пунктами і лісовими масивами;
- Нарешті, „острови“ міських лісів та/або лісопарків, які поступово переходять в приміські масиви рекреаційних лісів. Перші розділені не стільки вулицями тощо магістралями, скільки „плямами“ міської та промислової забудови, другі — в основному магістралями, меншою мірою „плямами“ дачної забудови, градієнт розділенності падає від центра урбанізованого „ядра“ до його зовнішньої межі і далі до периферії регіону.
Матрикс схожий з океаном, що розділяє острови, своєю непридатністю для життя відповідних видів, вони можуть перетинати його тільки транзитом. Відрізняється він від океана тим, що може показувати свого роду „напівпроникливості“, залежно від специфічної структури ландшафта, затримуючи одні види даної групи і сприяючи транзиту інших (при тому що всі відповідні види належать до однієї групи, екологічно і систематично близьких). Відповідно, антропогенно-перетворений ландшафт (і особливо міський) здійснює спрямований відбір видів до складу фауни „архіпелага“, сприяючи одним і перешкоджаючи іншим (далі ми побачимо, які характеристики життєвої стратегії виду визначають його попадання в першу і другу), чого аж ніяк не робить океан, що розділяє острови.
Виникає питання — чим відрізняється „архіпелаг“, створений антропогенною фрагментацією середовища існування, від природних систем островів? Головних якісних відмінностей дві:
- Антропогенний „архіпелаг“ перебуває в постійній динаміці, оскільки освоєння території, що створило його, не припиняється, а діє як мінімум на тому ж рівні або розвивається далі. Інша істотна відмінність „архіпелагів“, створених і підтримуваних антропогенної фрагментацією середовища існування, від справжніх острівних систем — це те, що вони не стабільні, а перебувають в постійному розвитку.
Зазвичай це пов'язане з чітким просторовим градієнтом, один полюс якого звернений до залишків первісного „материка“, інший знаходиться в області найбільшого антропогенного освоєння, порушення і трансформації ландшафту. У разі урбанізаційних процесів це градієнт „центр — периферія“ регіону, уздовж якого і розвивається „архіпелаг“.
Внутрішній компонент цієї динаміки полягає в зростаючому „дробленні“, ізоляції „островів“, зі стохастичним їх розміщенням один щодо одного. Тому в переважній більшості цих архіпелагів найбільші „острови“ одночасно зустрічаються групами, члени якої невипадково зближені між собою, найдрібніші — поодинці і рівновіддалені один від одного, з поступовим переходом з першого стану в другий у розвитку „архіпелага“.
Зовнішній полягає в тому, що одночасно з розвитком „архіпелага“ відбувається поступове „підгризання“ материка. Масиви останнього перетворюються на „острови“, що ізолюються і вбудовуються в існуючий патерн просторової конфігурації „архіпелагу“. Протилежний процес йде з протилежного полюса „архіпелага“. Там розвивається мережа техногенних аналогів природних місць існування (ставки рибгоспів як заміна водно-болотних угідь, старі парки — лісів, пустирі з різнотравними газонами — лук), що частково компенсують „дроблення“ з переварюванням» «островів», і різні "дикі"види намагаються туди «пересісти».
- У випадку справжніх островів кордони острова зазвичай спільні для всіх видів даної групи — птахів, метеликів, тощо У разі ж лісових мікрофрагментів в степу і вірних видів антропогенних «архіпелагів» часто виявляється, що різним видам птахів, метеликів або гризунів властиве різне «мереживо» місцеперебувань, що для одного виду буде «островом», для іншого «півостровом» або «материком» . У першу чергу це пов'язане з тим, що саме між близькими видами однієї гільдії існують максимальні відмінності за ступенем стабільності/нестабільності тих природних ландшафтів, в яких до початку впливу жили вони всі.
Коли господарське освоєння вносить «збурення», воно формує не тільки сам архіпелаг, але і задає рівень нестабільності на його «островах», зазвичай негативно пов'язаний з розміром «острова». Чим дрібніший «острів», тим швидше змінюються його обриси (а точніше, конфігурація «острівних» місцеперебувань в околиці, що включає сам «острів» і кільце того ж радіуса навколо нього часу, внаслідок змін рослинності). Включаючи появу/знищення нових аналогічних ділянок. Тому більш лабільні види, які миряться з більшою фрагментованістю/зміною місць існування, можуть формувати постійні поселення там, де їх близькі родичі з консервативнішими відносинами з територією, з біотопами — ні. Відповідно те, що для одного виду — один великий «острів», для іншого (екологічно близького, з того ж роду) — два дрібних ізоляти, з відповідними наслідками для стійкості популяції і конкретного «мережива ареалу» в «архіпелазі».
Формування острівних фаун
Заселення далеких островів полегшується при наявності між ними і материком проміжних острівців, так званих «кам'яних сходинок». Останні протягом багатьох геологічних відрізків часу могли зникати і знову з'являтися. За допомогою таких «сходинок» види могли розселятися на дуже великі відстані, використовуючи їх як тимчасове пристанище.
Зазвичай тварини, що розселяються з материка на архіпелаг, спочатку потрапляють на найближчі острови, а потім через вузькі протоки — на віддаленіші. Зрештою, на островах, розташованих уздовж міграційного шляху, створюється певна послідовність родинних форм, число яких у міру віддалення від материка поступово зменшується. Ф. Дарлінгтон називає такий тип розповсюдження імміграційним. У той же час на островах океанічного походження, за його твердженням, спостерігається й інший тип розповсюдження — реліктовий. При цьому на окремих островах якогось архіпелагу в результаті вимирання тих чи інших видів колишньої фауни з'являються «прогалини». На інших же островах даного архіпелагу ці види зберігаються. Тому послідовність в поширенні видів і родів на сусідніх островах порушується. Так як хвилі переселення видів з материка слідують одна за одною, на острові може виявитися змішана фауна, що складається з різновікових елементів.
У формуванні острівних фаун є ще одна важлива закономірність. Мова йде про закріплення переселенців на островах, інакше кажучи, про їх натуралізацію. Успіх натуралізації залежить від багатьох причин. Витриваліші й еврібіонтні види мають більше шансів на вкорінення, ніж стенобіонтні і маловитвалі. Якщо абіотичні умови острова значно відрізняються від умов тих місць, звідки прибули переселенці, їм доводиться пройти період акліматизації, в процесі якої можуть сильно змінитися їх екологічні особливості, а це може привести до формування нових підвидів і видів. Серйозно перешкоджає успішній натуралізації нових для острова видів конкуренція з боку тварин, що вже живуть на ньому. Між ними при схожості екологічних потреб починається запекла боротьба за існування. Природно, першопоселенці мають ряд переваг перед новоприбулими — у перших було більше часу для натуралізації, зокрема для збільшення чисельності своїх популяцій. Якщо ж нові види зуміли утриматися в складі фауни, то на кінцевому етапі колонізації настає взаємне пристосування тварин до існування в одному біоценозі.
Зазвичай острівні біоценози по видовій, просторовій і функціональній структурі значно простіше, ніж материкові. Як відомо, біоценози з простою структурою і бідним видовим складом дуже нестійкі і відрізняються слабкою буферністю. Цим і пояснюється вразливість острівних біоценозів при втручанні людини, будь це завезення нових для острова тварин, зміна місця існування або винищення будь-яких видів. Прикладів подібного роду багато.
Література
- Одум Ю. Основы экологии. — М.: Мир, 1975. — 740 с.
- Стольберг Ф. В. Экология города (урбоэкология). — К.: Либра, 2000. — 464 с.
- Яницкий О. Н. Экологическая перспектива города. — 1987.
- Sukopp, H. & Wittig, R. (Hrsg.) (1998): Stadtökologie. Ein Fachbuch für Studium und Praxis. 2. überarbeitete Auflage. — Stuttgart, Jena, Lübeck, Ulm (Gustav Fischer Verlag). — 474 S. ISBN 3-437-26000-6.
Ресурси Інтернета
Примітки
- MacArthur R.H., Wilson E.O. The theory of island biogeography. — Princeton: Princeton Univ. Press, 1967. — 293 p.
- Стольберг Ф. В. Экология города (урбоэкология). — К.: Либра, 2000. — 464 с.
- Яницкий О. Н. Экологическая перспектива города. — 1987.
- Sukopp, H. & Wittig, R. (Hrsg.) (1998): Stadtökologie. Ein Fachbuch für Studium und Praxis. 2. überarbeitete Auflage. — Stuttgart, Jena, Lübeck, Ulm (Gustav Fischer Verlag). — 474 S.
- Насимович Ю. А. Гидрографическая сеть Москвы. — С. 50-61.