Трансдукція (генетика)
Трансдукція (від лат. transductio — переміщення) — форма горизонтального перенесення генів, при якій передача генетичного матеріалу від однієї клітини до іншої відбувається за допомогою вірусу (бактеріофага у випадку бактерій), що, як і у випадку інших форм горизнотального перенесення генів, призводить до зміни спадкових властивостей. Вірус, що переносить клітинну ДНК або РНК, називається трансдукційною частинкою. Розрізняють два види трансдукції: загальну (генералізовану), за якої може переноситись будь-яка ділянка геному клітини, та спеціалізована, під час якої завжди переноситься один і той самий набір генів. Явище трансдукції було відкрите американськими вченими Джошуа Ледербергом і Нортоном Циндером у 1952 році, під час вивчення бактерії Salmonella typhimurium та її паразита фага P22. Явище загальної трансдукції використовують для картування геномів бактерій, а також у генній інженерії.
Історія дослідження
У 1952 році Циндер та Ледерберг спостерігали явище генетичного обміну між двома ауксотрофними штамами S.typhimurium, внаслідок якого утворювались прототрофні бактерії. Вони показали, що перенесення генів відбувається без фізичного контакту між клітинами, потрібного для кон'югації, і його проходженню не перешкоджає додавання ДНКази до середовища, а отже цей процес відмінний від трансформації. Агент, відповідальний за генетичний обмін був здатний проходити через бактерійні фільтри, і згодом було з'ясовано, що ним є помірний бактеріофаг P22. Це відкриття стало можливим через щасливий випадок: один із штамів, який використовували Ледерберг і Циндер, був лізогенізований бактеріофагом P22. На певній стадії експерименту в якійсь із клітин відбулась індукція профага, після чого сформовані вірусні частинки заразили інший ауксотрофний штам і перенесли до нього гени дикого типу від свого попереднього живителя[1].
Згодом аналогічний процес був виявлений і в інших гарм-негативних бактерій, зокрема Escherichia coli (здійснюється бактеріофагом P1), Myxobacteria, Rhizobium, Caulobacter та Psuedomonas. Як для фага P22, так і для фага P1, було показано, що трансдукція є результатом фізичного перенесення ДНК від донора до реципієнта: донорний штам вирощували у середовищі, яке містило важкі ізотопи Нітрогену та Гідрогену (2H та 15N), які включались у ДНК під час реплікації, тобто нові покоління бактерій містили «важку» ДНК. Через деякий час культуру переносили на звичайне середовище (із 1H та 14N) та інфікували бактеріофагами. В таких умовах синтезувалась «легка» вірусна ДНК. Було з'ясовано, що деякі вірусні частинки включають у свою структуру важку, тобто бактерійну, ДНК, коли такі бактеріофаги заражали реципієнтний штам, то у багатьох клітинах важка ДНК вбудовувалась у хромосому.[1]
Загальна трансдукція
Загальна трансдукція характеризується тим, що переноситись може будь-який фрагмент бактерійної хромосоми. Цей процес потребує успішного проходження двох етапів: утворення вірусної частинки, що містить ДНК реципієнта, та стабільне введення цієї ДНК у клітину донора, останнє найчастіше забезпечується сайт-специфічною рекомбінацією із бактерійною хромосомою.
Події в клітині донора
Запаковування бактерійної ДНК у вірусні голівки — це помилка у процесі реплікації вірусу. Вона може траплятись із різних причин, наприклад, якщо бактерійна ДНК містить послідовність дуже схожу до тієї, що слугує сигналом для початку пакування у фаговій ДНК. Загалом процес формування трансдукційних частинок залежить від особливостей метабіолізму ДНК конкретного виду вірусу. Найчастіше до трансдукції здатні фаги, пакування ДНК в яких відбувається за механізмом «повної голівки». Найкраще цей процес вивчений на прикладі фага P22.
Формування трансдукційних частинок фага P22
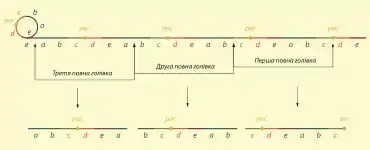
Геном фага P22 характеризується кінцевою надлишковістю (тобто крайня ліва послідовність, довжиною приблизно 2% від всієї ДНК, повторюється на правому кінці) та циклічною пермутацією (тобто ділянка, яка повторюється не є сталою, а відрізняється у ДНК кожної вірусної частинки). Такі особливості пов'язані пакуванням вірусної ДНК за механізмом «повної голівки» (англ. headful packaging).[1]
Вірусні частинки фага P22 містять лінійну дволанцюгову ДНК із «тупими» кінцями, тобто без однониткових виступів. Коли, після інфікування, вона потрапляє у клітину, то, в першу чергу, відбувається її циклізація. Перетворення лінійної ДНК у кільцеву здійснюється шляхом сайт-специфічної рекомбінації між правою і лівою кінцевими ділянками, і є можливою тільки завдяки тому, що ці ділянки однакові, в цьому і полягає значення існування кінцевої надлишковості у геномі фага P22. Далі кільцева ДНК вірусу реплікується за механізмом «кільця, що котиться», внаслідок чого утворюється довгий конкатамер, який складається із великої кількості послідовно сполучених копій геному фага.
Пакування ДНК у вірусні голівки починається із специфічної послідовності pac, тут фермент, що кодується вірусним геном, робить перший надріз, який визначає початок пакування. Довжина ДНК, яка буде запакована, визначається розміром голівки (звідси і назва механізму), коли остання наповнюється, фермент робить другий надріз. Цей надріз визначає не тільки закінчення формування першої голівки, а й початок формування другої. Тобто ДНК, що пакується у другий віріон вже не починається із pac-сайту і її початок не є специфічним. Третій надріз робиться, коли повністю запаковується друга голівка, після чого починається формування четвертого віріона. Отже під час пакування ДНК фага P22 специфічна послідовність необхідна тільки для ініціації процесу, далі він відбувається не залежно від якихось специфічних ділянок геному. Довжина ДНК, що запаковується визначається тільки розміром вірусної голівки, і у фага P22 є дещо більшою ніж загальна довжина геному, через що і виникає кінцева надлишковість, а оскільки надрізи щоразу робляться в іншій ділянці, то і повторюються в кожній утвореній молекулі ДНК різні фрагменти.[1]
Під час інфекції фагом P22 бактерійна хромосома не деградується, а отже вона також може виступати субстратом для пакування у вірусні частинки. В геномі S.typhimurium справді є 10—15 ділянок, схожих до pac-сайту фага P22, і в них може ініціюватись процес пакування. Таким чином утворюватимуться вірусні частинки, котрі нестимуть гени бактерії. Проте трансдукційні частинки формуються значно рідше ніж звичайні віріони, імовірно, через те, що хромосомні ділянки, з яких починається пакування бактерійної ДНК, не повністю ідентичні до pac-сайту вірусу. Починаючи із кожної з таких послідовностей у голівки може запаковуватись 10—15% геному бактерії, проте, чим дальше ген знаходиться від pac-сайту, тим менша імовірність його потрапляння у трансдукційну частинку. Через це деякі гени S.typhimurium переносяться фагом P22 у тисячу разів рідше, ніж інші.
Існує HT мутант фага P22, що характеризується значно більшою частотою формування трансдукційних частинок (приблизно половина вірусного потомства), ніж дикий тип. Імовірність перенесення деяких генів такими вірусами може збільшуватись до 10 000 разів порівняно із вихідним штамом. HT мутант має змінений білок, що розпізнає pac-сайт. Через зсув у специфічності він починає «віддавати перевагу» іншій, значно поширенішій в геномі бактерії, послідовності. HT штам часто використовують у певних генетичних маніпуляціях, для яких потрібна трансдукція[1].
Формування трансдукційних частинок іншими фагами
Механізм загальної трансдукції, з'ясований під час дослідження фага P22, виявився справедливим і для багатьох інших бактеріофагів. Наприклад, перенесення генетичної інформації фагом P1 E.coli відбувається за такою ж загальною схемою, які і у випадку P22, проте має певні відмінності: по-перше фагова голівка P1 приблизно вдвічі більша за голівку P22, а отже вдвічі більша і довжина ДНК, що запаковується, по-друге хромосома кишкової палички містить значно більше сайтів ініціації пакування (pac) ніж хромосома S.typhimurium. Через це коливання частоти трансдукції різних генів значно менші у випадку фага P1, порівняно із P22.
До загальної трансдукції здатні далеко не всі бактеріофаги, наприклад, вірулентний фаг T4 E.coli спричинює деградацію бактерійної ДНК, а отже у голівки може пакуватись тільки його власна. ДНК цього вірусу містить модифікований нуклеотид — глікозильований гідроксиметилцитозин. Бактерійна хромосома руйнується фаговою нуклеазою, що діє тільки на ДНК із немодифікованим цитозином, в той час як вірусна ДНК залишається неушкодженою. У 1979 році був отриманий мутант фага T4 із дефектною системою дегарадації хромосоми живителя, цей штам здатний ефективно здійснювати загальну трансдукцію.
Для фага λ характерна здатність до спеціалізованої, а не загальної, трансдукції. Пакування ДНК цього вірусу відбувається не за механізмом «повної голівки», а є сайт-специфічним. Під час пакування кожен надріз робиться тільки у cos-сайті, при чому формуються «липкі кінці». Проте було встановлено, що на 60—90 хвилині літичного розвитку фага лямбда стає можливим запаковування бактерійної ДНК у фагові голівки без розпізнавання cos-сайтів. Цей процес пригнічується певною вірусною екзонуклеазою. Мутанти, у яких ця екзонуклеаза «вимкнена», як і гени, відповідальні за лізис, можуть використовуватись як ефективні агенти для загальної трансдукції.
Події в клітині реципієнта
Трансдукційні частинки, як і звичайні бактеріофаги, здатні успішно вводити ДНК у бактерійні клітини, при цьому інфекція не розвивається, оскільки гени відповідальні за реплікацію вірусу відсутні. ДНК донора в клітині реципієнта може або вбудовуватись у хромосому, або залишатись в цитоплазмі. В останньому випадку, якщо даний фрагмент ДНК не здатний до реплікації, він буде з часом втрачений (абортивна трансдукція), якщо ж він може реплікуватись автономно, то передаватиметься в наступні покоління як позахромосомний носій спадковості.
Вбудовування донорної ДНК у ДНК реципієнта відбувається шляхом гомологічної рекомбінації. Принаймні у випадку фагів P22 та P1 відбувається двониткова заміна фрагменту бактерійної хромосоми на введену вірусом ДНК, для цього необхідне проходження двох кросинговерів, по одному на кожному із кінців ділянки, що вбудовується. Якщо ці кросинговери відбуватимуться посередині інтродукованого фрагмента ДНК, то включатиметься в хромосому відповідна лише його частина. Із всієї ДНК, що переноситься в клітину-реципієнта під час трансдукції в кінцевому результаті приблизно лише 5% опиняється в її хромосомі.
Використання загальної трансдукції
Загальну трансдукцію широко використовують для побудови генетичних карт, тобто для встановлення послідовності розташування генів, а також відстані між ними. Для цього використовують котрансдукцію двох або частіше трьох факторів. Зазвичай таким методом досліджують гени, розташовані надто близько, щоб їх послідовність можна було встановити методом перерваної кон'югації.
| Клас | Кількість |
|---|---|
| A+B+C+ | 50 |
| A+B+C- | 75 |
| A+B-C- | 300 |
| A+B-C+ | 1 |
Наприклад, якщо потрібно картувати три близьких гени A, B і C як донор використовуватиметься прототроф A+B+C+, на культурі якого вирощуватимуться бактеріофаги, частина з яких утворить трансдукційні частинки. Отриманими фагами заражатимуть ауксотрофний штам A-B-C-, при цьому утвориться сім класів бактерій, у яких відбулась трансдукція, і один, найбільший, в якому не відбулось, останнього слід позбутись для подальших досліджень. Найпростіший метод — це відібрати клітини, в яких ген дикого типу замінив хоча б один із локусів, для прикладу, нехай це буде ген A. Культуру бактерій вирощуватимуть на мінімальному середовищі, із доданими факторами, необхідними для ауксотрофів B- та C-, таким чином можна відібрати чотири класи трансдуктантів A+B-C-, A+B+C-, A+B-C+, A+B+C+, ще три будуть втрачені. Після цього репліки отриманих культур переносять на повноцінне середовище без фактору, необхідного для B- та на повноцінне середовище без фактору для C-, таким чином можна визначити, чи відбулась трансдукція у кожному з цих двох локусів. Тепер можна обрахувати частоту кожного із чотирьох класів, дані для прикладу наведені у таблиці справа. З отриманих результатів можна зробити висновок, що ген B знаходиться між генами A і С, оскільки найрідкіснішим завжди є той, клас, в якого в хромосому вбудувались два крайніх гени, але не середній, через те, що в такому випадку повинно було відбутись не два, а чотири кросинговери. Проте, результати, отримані таким методом будуть хоч і інформативними але неповними, через втрату трьох класів трансдуктантів.[2]
Справжня залежність чатсоти котрансдукції від відстані між генами є нелінійною, оскільки на неї впливають два різних фактори: по-перше, чим ближче розташовані два гени, тим рідше кросинговер, потрібний для вбудовування ДНК донора у ДНК реципієнта, проходитиме між ними, по-друге, відстань також впливатиме і на ймовірність запаковування цих генів в одну фагову голівку[1].
Крім картування генів загальна трансдукція також може використовуватись для комплементаційного аналізу, перенесення плазмід та транспозонів, конструюювання нових штамів мікроорганізмів[1].
Спеціалізована трансдукція
Спеціалізована трансдукція відрізняється від загальної кількома рисами: по-перше, під час цього процесу ділянка ДНК донора у трансдукційній частинці ковалентно приєднана до вірусної ДНК, по-друге переносяться не будь-які гени, а лише певна специфічна ділянка бактерійної хромосоми. Спеціалізована трансдукція використовуються як потужний інструмент для маніпуляції генами: молекулярного клонування, картування окремих генів та мутацій в них, спрямованого мутагенезу, комплементаційного аналізу тощо[1].
Формування трансдукційних частинок фага лябмда
Найкраще вивченим прикладом спеціалізованої трансдукції є трансдукція за допомогою помірного бактеріофага λ E.coli. Геном цього вірусу представлений лінійною двонитковою ДНК із 12-нуклеотидними «липкими кінцями». Потрапляючи в клітину бактерії ця ДНК замикається у кільце завдяки гібридизації між «липкпими кінцями», на місця яких формується cos-сайт. На певному етапі літичного циклу розвитку фага λ його геном реплікуються за механізмом «кільця, що котиться», внаслідок чого утворюються довгі конкатамери. Під час пакування ДНК у фагові голівки, ферменти, що розрізають конкатамер на окремі фрагменти, діють сайт-специфічно — тільки у cos-ділянках, генеруючи «липкі кінці». Довжина ДНК, що пакується може бути різною — від 35 до 50 т.п.н.[1].
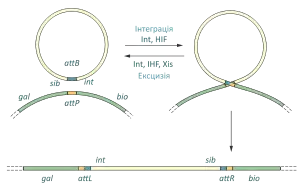
Під час лізогенного циклу розвитку геном фага λ вбудовується у хромосому кишкової палички між галактозним та біотиновим опероном, завдяки рекомбінації між ділянками aatP (англ. phage attachment) та aatB (англ. bacteria attachment), при цьому на кінцях вставки (профага) утворюються дві нові ділянки attR (англ. attachment site on the right) та attL (англ. attachment site on the left). В разі індукції профага, він повинен вирізатись із хромосоми кишкової палички. Зазвичай цей процес відбувається шляхом рекомбінації між сайтами attR та attL, проте інколи відбувається помилкова, так звана «незаконна», рекомбінація, у випадку якої фрагмент, що вирізається зсувається вправо або вліво, а отже він міститиме частину бактерійної ДНК і не міститиме частини вірусної, якщо при цьому cos-сайт не втарчається, то така ДНК може успішно запаковуватись у вірусні голівки. Переважно трансдукційні частинки фага лямбда переносять галактозний оперон E.coli, такі фаги позначаються λ gal. Частина із цих вірусів несуть всі необхідні для літичного розвитку гени, інші є дефектними і можуть розвиватись тільки у присутності фага-помічника[1].
Події в клітині реципієнта
Коли фаг, що несе ділянку хромосоми донора, заражає клітину реципієнта стабільна трансдукція може відбуватись кількома шляхами. Фаг може лізогенізувати клітину, якщо у нього зберігся сайт attB і ген int, то вбудовування відбувається аналогічно як і в дикого типу. Проте, зазвичай якраз ці ділянки і втрачаються у процесі формування трансдукційної частинки, тому вбудовування відбувається іншим чином, найчастіше шляхом рекомбінації між ділянкою ДНК донора, принесеною фагом, та гомологічною ділянкою в хромосомі живителя. За деяких умов геном трансдукційного вірусу може зберігатись в цитоплазмі клітини як плазміда[1].
Абортивна трансдукція
Абортивна трансдукція — трансдукція, при якій ділянка хромосоми бактерії, що потрапляє в іншу бактеріальну клітину, не подвоюється при її наступних поділах, а передається тільки одній дочірній клітині.[3]
Джерела
- Streips UN, Yasbin RE (2002). Modern Microbial Genetics (вид. 2nd). Willey-Liss, Inc. ISBN 0471386650.
- Tamarin RH (2001). Principles of Genetics (вид. 7th). Mcgraw-Hill. ISBN 0072334193.
- Абортивная трансдукция // Большая советская энциклопедия / главн. ред. А. М. Прохоров. — 3-е изд. — Тома 1–30. — М.: «Советская энциклопедия», 1969–1978. (рос.).
Посилання
- Трансдукція // Універсальний словник-енциклопедія. — 4-те вид. — К. : Тека, 2006.
