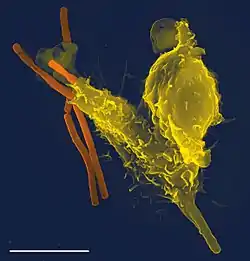
Імунна система
Іму́нна систе́ма — сукупність органів, тканин, клітин, які забезпечують захист організму від чужорідних агентів; система організму, яка контролює сталість клітинного і гуморального складу організму.
Знищенню імунною системою підлягає генетично чужорідне: молекули інших організмів, мікробні клітини, молекули, до яких утворюються антитіла, а також, пошкоджені клітини власного організму; крім того, імунна система може реагувати на власні клітини та тканини, що мають пошкодження або злоякісно трансформовані.
| Імунна система | |
|---|---|
 | |
| Ідентифікатори | |
| MeSH | D007107 |
| Анатомічна термінологія | |
Історія розвитку уявлень про імунну систему
У 1796 році англійський сільський лікар Едвард Дженнер (1749—1823) розробив спосіб штучної імунізації проти натуральної віспи шляхом зараження людини коров'ячою віспою.
Відкриття Луї Пастера (1880) дало початок імунології як самостійній науці. Пастер виявив, що імунізація курей старою холерною культурою створює у них стійкість до зараження високовірулентним збудником курячої холери і сформулював основний принцип створення вакцин, а також отримав їх проти сибірки і сказу.
Ілля Ілліч Мечников відкрив феномен фагоцитозу (1882) і створив клітинну або фагоцитарну теорію імунітету.
Німецький учений Пауль Ерліх висунув гуморальну теорію імунітету. З 1898 по 1899 рік бельгійський науковець Жюль Борде і російський М. Я. Чистович виявили утворення антитіл у відповідь на введення чужорідних еритроцитів і сироваткових білків. Дане відкриття поклало початок неінфекційній імунології.
У 1900 році австрійський імунолог Карл Ландштейнер зробив відкриття груп крові AB0 людини[джерело?]. Він створив основу вчення про тканинні ізоантигени.
Органи імунної системи
- центральні (кістковий мозок і тимус)
- периферійні (селезінка, лімфатичні вузли та інші накопичення лімфоїдних тканин)
Клітини імунної системи
- лейкоцити (спеціальні клітини імунної системи):
- лімфоцити (Т-лімфоцити, В-лімфоцити, Нормальні кілери)
- фагоцити (макрофаги, еозинофіли, нейтрофіли, базофіли, дендритні клітини, мікроглії, купферовські клітини)
- допоміжні клітини (тучні клітини, тромбоцити)
Лімфоцити
Клітини імунної системи, на які покладені ключові функції щодо здійснення набутого імунітету, належать до лімфоцитів, які є підтипом лейкоцитів. Велика частина лімфоцитів відповідає за специфічний набутий імунітет, оскільки можуть розпізнавати збудників інфекції всередині або поза клітинами, в тканинах або в крові.
Основними типами лімфоцитів є B-лімфоцити та T-лімфоцити, які формуються з плюріпотентних гемопоетичних стовбурових клітин; у дорослої людини вони утворюються в кістковому мозку, а T-лімфоцити додатково проходять частину етапів диференціювання в тимусі. B-клітини відповідають за гуморальну ланку набутого імунітету, тобто виробляють антитіло, в той час як T-клітини є основою клітинної ланки специфічної імунної відповіді.
На етапі розвитку лімфоцити поділяються на групи, здатні виконувати ту чи іншу функцію захисту. Існують різні види лімфоцитів. Зокрема, за морфологічними ознаками їх поділяють на малі лімфоцити та великі гранулярні лімфоцити (ВГЛ). За структурою зовнішніх рецепторів серед лімфоцитів виділяють, зокрема, B-лімфоцити та T-лімфоцити.
Як B-, так і T-клітини несуть на своїй поверхні рецепторні молекули, які розпізнають специфічні мішені. Рецептори являють собою як би «дзеркальний відбиток» певної частини чужорідної молекули, здатний приєднуватися до неї. При цьому одна клітина може містити рецептори тільки для одного виду антигенів.
T-клітини розпізнають чужорідні («не-свої») мішені, такі як патогенні мікроорганізми, тільки після того, як антигени (специфічні молекули чужорідного тіла) будуть оброблені та презентовані в поєднанні з власною молекулою головного комплексу гістосумісності (англ. main histocompatibility complex, MHC). Серед T-клітин розрізняють ряд підтипів, зокрема, Т-кілери, Т-хелпери та регуляторні Т-клітини.
T-кілери розпізнають тільки антигени, які об'єднані з молекулами головного комплексу гістосумісності I класу, у той час як T-хелпери розпізнають тільки антигени, розташовані на поверхні клітин в поєднанні з молекулами головного комплексу гістосумісності II класу. Ця різниця у презентації антигену відображає різні ролі зазначених двох типів T-клітин. Іншим, менш поширеним підтипом T-клітин, є γδ T-клітини, які розпізнають незмінені антигени, не пов'язані з рецепторами головного комплексу гістосумісності[1].
У T-лімфоцитів коло завдань достатньо широке. Частина з них — регуляція набутого імунітету за допомогою спеціальних білків (зокрема, цитокінів), активація B-лімфоцитів для утворення антитіл, а також регуляція активації фагоцитів для ефективнішого руйнування мікроорганізмів. Цю задачу виконує група T-хелперів. За руйнування власних клітин організму шляхом виділення цитотоксичних факторів при безпосередньому контакті відповідають T-кілери, які діють специфічно.
На відміну від T-клітин, B-клітини не потребують обробки антигену та експресії його на поверхні клітини. Їх рецептори до антигену є фіксовані на поверхні B-клітини антитілоподібні білки. Кожна лінія B-клітин, що пройшла диференціювання, експресує унікальне тільки для неї антитіло, і ніяке інше. Таким чином, повний набір антигенних рецепторів всіх B-клітин організму представляє всі антитіла, які організм може виробляти. Функція B-лімфоцитів полягає насамперед у виробленні антитіл — гуморального субстрату специфічного імунітету —, дія яких спрямована насамперед проти позаклітинних збудників.
Крім того, існують лімфоцити, які не специфічно проявляють цитотоксичність — природні кілери.
T-кілери
Т-кілери являють собою підгрупу T-клітин, функцією яких є руйнування власних клітин організму, інфікованих вірусами або іншими патогенними внутрішньоклітинними мікроорганізмами[2], або клітин, які пошкоджені або невірно функціонують (наприклад, пухлинні клітини). Як і B-клітини, кожна конкретна лінія T-клітин розпізнає тільки один антиген. T-кілери активуються при з'єднанні своїм T-клітинним рецептором (ТКР) зі специфічним антигеном в комплексі з рецептором головного комплексу гістосумісності I класу іншої клітини. Розпізнавання цього комплексу рецептора гістосумісності з антигеном здійснюється за участю розташованого на поверхні T-клітини допоміжного рецептора CD8. У лабораторних умовах T-клітини зазвичай виявляють саме по експресії CD8. Після активації T-клітина переміщується по організму в пошуках клітин, на яких білок I класу головного комплексу гістосумісності містить послідовність потрібного антигену. При контакті активованого T-кілера з такими клітинами він виділяє токсини, що утворюють отвори в цитоплазматичній мембрані клітин-мішеней, в результаті іони, вода та токсин вільно переміщуються в клітку-мішень та з неї: клітина-мішень гине[3]. Руйнування власних клітин T-кілерами важливо, зокрема, для запобігання розмноженню вірусів. Активація T-кілерів жорстко управляється та "зазвичай вимагає дуже сильного сигналу активації від комплексу білка гістосумісності з антигеном, або додаткової активації факторами T-хелперів[3].
T-хелпери
Т-хелпери регулюють реакції як вродженого, так і набутого імунітету, і дозволяють визначати тип відповіді, який організм зробить на конкретний чужорідний матеріал[4][5]. Ці клітини не виявляють цитотоксичності та не беруть участь у знищенні інфікованих клітин або безпосередньо збудників. Замість цього, вони керують імунною відповіддю, спрямовуючи інші клітини на виконання цих завдань.
T-хелпери експресують T-клітинні рецептори (ТКР), які розпізнають антигени, пов'язані з молекулами II класу головного комплексу гістосумісності. Комплекс молекули головного комплексу гістосумісності з антигеном також розпізнається корецептором клітин-хелперів CD4, який привертає внутрішньоклітинні молекули T-клітини (наприклад, LCK), відповідальні за активацію T-клітини. T-хелпери мають меншу чутливість до комплексу молекули головного комплексу гістосумісності та антигену, ніж T-кілери, тобто для активації T-хелперів потрібне зв'язування набагато більшої кількості його рецепторів (близько 200—300) з комплексом молекули гістосумісності та антигену, в той час як T-кілери можуть бути активовані після зв'язування з одним таким комплексом. Активація T-хелперів також вимагає тривалішого контакту з антиген-презентируючою клітиною[6]. Активація T-хелперів призводить до вивільнення ними цитокінів, які впливають на активність багатьох видів клітин. Цитокінові сигнали, створювані T-хелперами, посилюють бактерицидну функцію макрофагів та активність T-кілерів[7]. Крім того, активація T-хелперів викликає зміни в експресії молекул на поверхні T-клітини, зокрема ліганду CD40 (також відомого під позначенням CD154), що створює додаткові стимулюючі сигнали, зазвичай необхідні для активації вироблення антитіл B-клітинами[8].
Гамма-дельта T-клітини
5-10 % T-клітин несуть на своїй поверхні ТКРгамма-дельта та позначаються як γδ T-клітини.
B-лімфоцити та антитіла
В-клітини становлять 5-15 % циркулюючих лімфоцитів та характеризуються поверхневими імуноглобулінами, вбудованими в клітинну мембрану та виконують функцію специфічного антигенного рецептора. Цей рецептор, специфічний лише для певного антигену, називається антитілом. Антиген, зв'язуючись з відповідним антитілом на поверхні В-клітини, індукує проліферацію та диференціювання В-клітини до плазматичних клітин та клітин пам'яті, специфічність яких така ж, як і специфічність вихідної В-клітини. Плазматичні клітини секретують велику кількість антитіл у вигляді розчинних молекул, які розпізнають вихідний антиген. Секретовані антитіла мають ту ж специфічність, що й відповідний В-клітинний рецептор.
Антиген-презентуючі клітини
Функція
Імунна система розпізнає чужорідні речовини, нейтралізує їх і «запам'ятовує» свою відповідь, щоб відтворити її при зіткненні з аналогічним антигеном. До компетенції імунної системи відносять і знищення клітин і білків власного організму, що виникають при нормальному, фізіологічному функціонуванні організму в екстремальних умовах — при травмах. В будь-якому стані організму імунна система постійно працює, хоч і з різним ступенем активності.
Імунітет
| Вроджений імунітет | Набутий імунітет |
|---|---|
| Реакція неспецифічна | Специфічна реакція, прив'язана до чужорідного антигену |
| Зіткнення з інфекцією призводить до негайної максимальної реакції | Між контактом з інфекцією і максимальною відповіддю латентний період |
| Клітинні і гуморальні ланки | Клітинні і гуморальні ланки |
| Не має імунологічної пам'яті | Зіткнення з чужорідним агентом призводить до імунологічної пам'яті |
| Виявляється практично у всіх форм життя | Виявлено тільки у деяких організмів |
Набутий імунітет
Система набутого імунітету з'явилася в ході еволюції нижчих хребетних. Вона забезпечує інтенсивнішу імунну відповідь, а також імунологічну пам'ять, завдяки якій кожний чужорідний мікроорганізм або чужорідна молекула «запам'ятовується» за унікальними для нього антигенами[9]. Система набутого імунітету антигеноспецифічна та вимагає розпізнавання специфічних антигенів в процесі, що має назву презентація антигена. Специфічність антигену дозволяє здійснювати реакції, які призначені конкретним мікроорганізмам або інфікованим ними клітинам. Здатність до здійснення таких вузьконаправлених реакцій підтримується в організмі «клітинами пам'яті». Якщо макроорганізм інфікується мікроорганізмом більше одного разу, ці специфічні клітини пам'яті використовуються для швидкого знищення такого мікроорганізму.
Імунологічні розлади у людини
Імунодефіцити
Імунодефіцити (ІДС) — це порушення імунологічної реактивності, які зумовлені випаданням одного або декількох компонентів імунного апарату або тісно взаємодіючих з ним неспецифічних факторів.
Автоімунні процеси
Автоімунні процеси — це значною мірою хронічні явища, які призводять до довготривалого пошкодження тканин. Це пов'язано в першу чергу з тим, що автоімунна реакція постійно підтримується тканинними антигенами.
Гіперчутливість
Гіперчутливість — це термін, використовуваний для позначення імунної відповіді, яка протікає в агравірованій і неадекватній формі, в результаті чого відбувається пошкодження тканин.
Алергія є передумовою виникнення патологічного імунологічного процесу. Алергени — речовини, які спричиняють алергічні реакції. Є зовнішні алергени (екзоалергени): харчові продукти, хімічні речовини, лікарські препарати, запахи). Внутрішні алергени (ендоалергени) — це власні тканини організму переважно з видозміненими властивостями, що виникають внаслідок утворення токсичних речовин при патологічних процесах (опіки чи обмороження, дія отруйних речовин, укуси бджіл, іонізуюча радіація).
Див. також
Примітки
- Holtmeier W, Kabelitz D. gammadelta T cells link innate and adaptive immune responses. Chem Immunol Allergy 86: 151–83. PMID 15976493.
- Harty J, Tvinnereim A, White D. CD8+ T cell effector mechanisms in resistance to infection. Annu Rev Immunol 18: 275–308. PMID 10837060.
- Radoja S, Frey A, Vukmanovic S (2006). T-cell receptor signaling events triggering granule exocytosis. Crit Rev Immunol 26 (3): 265–90. PMID 16928189.
- Abbas A, Murphy K, Sher A (1996). Functional diversity of helper T lymphocytes. Nature 383 (6603): 787–93. PMID 8893001.
- McHeyzer-Williams L, Malherbe L, McHeyzer-Williams M. Helper T cell-regulated B cell immunity. Curr Top Microbiol Immunol 311: 59–83. PMID 17048705.
- Kovacs B, Maus M, Riley J, Derimanov G, Koretzky G, June C, Finkel T (2002). Human CD8+ T cells do not require the polarization of lipid rafts for activation and proliferation. Proc Natl Acad Sci U S A 99 (23): 15006–11. PMID 12419850.
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1.
- Grewal I, Flavell R. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol 16: 111–35. PMID 9597126.
- Pancer Z, Cooper M. The evolution of adaptive immunity. Annu Rev Immunol 24: 497–518. PMID 16551257.
Джерела
- Ройт А., Брестофф Дж., Мейл Д. «Иммунология» / Пер. с англ. — М.: «Мир», 2000. — 592 с. (рос.)
- Иммунная система: центральные и периферические органы (рос.)
- Клетки, осуществляющие иммунный ответ: введение (рос.)