Рецептор (білок)
Рецептор — це велика білкова молекула, яка специфічно реагує зміною своєї просторової конформації на приєднання до неї молекули хімічної речовини (ліганд) або на фізичний стимул (механічний, тепловий, електричний тощо). Такі білки знаходяться на поверхні клітинної мембрани, клітинної органели або в цитоплазмі клітини. Рецепторний білок передає сигнал, який призводить до зміни функції клітини або її певної частини. Механізми рецепторної відповіді складають активацію вторинних месенджерів, генерацію електричного потенціалу в мембрані нейронів, зміну експресії генів
Хімічні рецептори
Принципи хімічної рецепції
Хімічна речовина, що специфічно з'єднується з деяким рецептором, називається лігандом цього рецептору.
Загальним правилом є те, що якщо в клітині знайдено рецептор для зв'язування деякої речовини штучного походження, то має існувати і природне з'єднання, з яким зв'язується даний рецептор (в більшості випадків така речовина виробляється в організмі, де знайдений рецептор) — так званий ендогенний ліганд. Наприклад, ендогенними лігандами опіатних рецепторів, зв'язування з якими обумовлює специфічний ефект похідних опійного маку та синтетичного наркотику метадону, є речовини білкової природи — ендорфіни. Ендогенними лігандами бензодіазепінового сайту (місця зв'язування) ГАМКА-рецептору, з яким зв'язуються синтетичні транквілізатори (такі як феназепам та діазепам) є особливі білки — ендозепіни. Ендогенним лігандом канабіноїдних рецепторів, з якими зв'язуються алкалоїди коноплі, є речовина анандамід, яка виробляється в організмі з арахідонової кислоти.
Будова рецептору
Окрім основної ділянки, або сайта зв'язування, де зв'язується специфічний ліганд, у рецептору можуть бути наявні також і додаткові ділянки, з якими зв'язуються речовини, що змінюють (модулюють) реакцію рецептору на основний ліганд — речовини-модулятори. Такі модулятори можуть посилювати або послаблювати реакцію на основний ліганд, або навіть замінювати собою його. Класичним прикладом такого рецептору з декількома ділянками зв'язування є ГАМКА-рецептор. Окрім сайта зв'язування ГАМК на ньому є ділянка, до якої приєднуються бензодіазепінові транквілізатори, сайт зв'язування барбітуратів, сайт зв'язування нейростероїдів і т. ін.
Багато рецепторів, залежно від речовини, що до них приєднується, можуть перебувати не тільки в конформаціях (або просторових концігураціях) «генерації сигналу» та «відсутності сигналу», а й декількох проміжних — тобто генерувати сигнали різної інтенсивності.
Ліганди
Речовина, що із 100% вірогідністю викликає перехід рецептору в конфігурацію «100% сигналу» називається повним агоністом рецептору. Наприклад, для ГАМКА-рецептору повним агоністом є діазепам. Речовина, що із 100% вірогідністю викликає перехід рецептору в конфігурацію «100% відсутності сигналу», називається повним антагоністом рецептору. Наприклад, для гліцинового рецептору повним антагоністом є стрихнін. Речовина, що викликає перехід рецептору в одну з проміжних стадій, або така, що викликає стан «100% сигналу» чи «100% відсутності сигналу», але з вірогідністю <100% (тобто частина рецептрів при зв'язуванні з цим лігандом перейде в одну з означених конформацій, а частина — ні), називається частковим агоністом або частковим антагоністом (залежно від переваги агоністичної або антагоністичної активності в його ефекті). Наприклад, синтетичний опіоїд бупренорфін є частковим агоністом μ-опіоідних рецепторів та частковим антагоністом κ-опіоїдних рецепторів.
Речовина, що зв'язується з тим самим сайтом на рецепторі, що і його ендогенний ліганд, називається такою, що завдає конкурентний ефект. Залежно від спрямованості ефекту це може бути конкурентний агоніст або конкурентний антагоніст (в обох випадках повний або частковий). Наприклад, бікукуллін є повним конкурентним антагоністом ГАМКА-рецептору, а (2S, 4R)-4-метілглутамінова кислота є конкурентним агоністом рецепторів каінової кислоти (підтип глутаматних рецепторів). Речовина, що змінює ефект рецептору, але при цьому зв'язується не з тим сайтом, з яким зв'язується ендогенний ліганд, називається (залежно від спрямованості дії) алостеричним (або неконкурентним) агоністом або антагоністом. Наприклад, барбітурати є алостеричними агоністами ГАМКА-рецептору, а речовина МК(+)801 є алостеричним антагоністом NMDA-рецепторів. Схематичне зображення 5-HT3-рецептора.
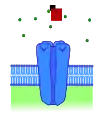 Рецептор без лігандів
Рецептор без лігандів Рецептор, активований ендогенним лігандом
Рецептор, активований ендогенним лігандом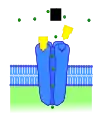 Рецептор, активований іншим конкурентним агоністом
Рецептор, активований іншим конкурентним агоністом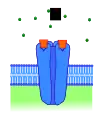 Рецептор, заблокований конкурентним антагоністом
Рецептор, заблокований конкурентним антагоністом
Механізми передачі сигналу рецепторами
Зміна конформації молекули рецептору призводить до запуску каскадів молекулярних подій. Білок-рецептор активується і починає виконувати функцію сам або ж активує інші молекули, які передають сигнал до кінцевого ефектору. Цим ефектором може бути інший білок, який починає виконувати нову функцію, або геном клітини, який змінює свою активність і впливає на роботу ансамблів білків. Чим далі від рецептору знаходиться ефектор, тим довший молекулярний каскад може бути задіяний.
Системи вторинних месенджерів
У відповідь на приєднання ліганду чи фізичний стимул багато мембранних рецепторів ініціюють синтез або деградацію внутрішньоклітинних малих молекул, що зв'язуються із цитоплазматичними рецепторами. Внутрішньоклітинні ліганди називають вторинними посередниками або месенджерами (на відміну від первинних посередників — позаклітинних). Вторинні посередники найчастіше здійснюють функцію через активацію протеїнкіназ та фосфатаз, які через механізм фосфорилювання білків змінюють їх активність.
Основним класом мембранних білків є рецептори, зв'язані з G-білками. G-білки активують або пригнічують роботу аденілатциклази, гуанілатциклази та фосфоліпази C, впливаючи на синтез вторинних посередників: цАМФ, цГМФ, інозитол-3-фосфату (І3Ф), діацилгліцеролу (ДАГ). Також вторинним посередником можуть виступати іони кальцію, що входять через кальцієві канали плазматичної мембрани або через І3Ф-активовані кальцієві канали ендоплазматичного ретикулуму, зв'язуються з кальмодуліном та за його посередництва активують протеїнкінази, білки м'язового скорочення тощо.
Електрогенні рецептори
Значна кількість рецепторів, особливо у постсинаптичних мембранах нервових і м'язових клітин, при з'єднанні з лігандом відкриває або закриває ворота каналу для проходження іонного струму. Струм може викликати деполяризувати мембрану, збуджуючи клітину, гіперполяризувати для процесів гальмування, а також кальцій, що входить через такі рецепторні канали, може виступати як вторинний посередник.
Ядерні рецептори
Цитоплазматичні та ядерні рецептори після утворення комплексу ліганд-рецептор мігрують до хроматину ядра, де виконують роль факторів транскрипції.
Різноманіття рецепторів
Існує декілька сотень генів, що кодують різноманітні рецепторні білки. Ці гени експресуються як у спеціалізованих клітинах органів чуттів і нервової системи, так і в усіх інших незбудливих клітинах.
Рецептори — іонні канали
- ГАМКА-рецептор
- ГАМКС-рецептор
- Нікотиновий ацетілхоліновий рецептор
- Гліциновий рецептор
- Іонотропні глутаматні рецептори
- Серотоніновий 5-HT3-рецептор
Рецептори, сполучені з G-білками
- Мускариновий ацетілхоліновий рецептор
- ГАМКВ-рецептор
- Адренергічні рецептори
- Дофамінові рецептори
- Метаботропні глутаматні рецептори
- Опіоїдні рецептори
- Гістамінові рецептори
- Брадикінінові рецептори
- Серотонінові рецептори
- Канабіноїдні рецептори
Рецептори мітогенів та факторів росту
- Інсуліновий рецептор
- Соматомедінові рецептори
- Рецептор фактору росту епідермісу
- Рецептор передсердного Na-діуретичного пептиду (NPR1, NPR2, NPR3)
Внутрішньоклітинні рецептори гормонів
- Глюкокортикоїдні рецептори
- Мінералокортикоїдні рецептори
- Естрогенові рецептори
- Гестагенові рецептори
- Андрогенові рецептори
- Тиреоїдні рецептори
Терморецептори
В основному електрогенні рецептори, що активуються як хімічними лігандами, так і зміною температури.
Механорецептори
Іонні канали, що відкриваються у вілповідь на деформацію мембрани.
- двопородоменні калієві канали
- протончутливі ASIC
- низькопороговий кальцієвий канал Cav3.2
- Канали транзієнтного рецепторного потенціалу, зокрема, TRPA1
- Piezo-рецептори, зокрема PIEZO1 та PIEZO2
Рецептори кислотності
Деякі йонні канали активуються іонами водню.
- епітеліальні ENaC
- протончутливі ASIC
Фоторецептори
- опсини
Див. також
- Афінітет
Джерела
- Ганонг, Вільям Ф. (2002). Фізіологія людини (Переклад з англ.). Львів: БаК. с. 784. ISBN 0-8385-8282-6.
