Джгутик
Джгутик (лат. flagellum) — рухома структура, присутня на поверхні клітин багатьох прокаріотів і еукаріотів. Служить для пересування в рідкому середовищі або поверхнею вологих твердих середовищ. Джгутики прокаріотів і еукаріотів різко відрізняються за своєю будовою: бактерільний джгутик має товщину 10-20 нм і довжину 3-15 μм, він пасивно обертається розташованим в мембрані мотором; тоді як джгутики еукаріотів мають товщину до 200 нм і довжину до 200 мкм, вони можуть самостійно згинатися на всій довжині. У еукаріотів часто також присутні війки, ідентичні за своєю будовою джгутику, але коротші (до 10 мкм); разом вони називаються ундуліподіями.
Джгутики бактерій
Джгутики бактерій складаються з трьох субструктур:
- Філамент (фібрила, пропелер) — порожнисте білкове волокно товщиною 10-20 нм і довжиною 3-15 μм, що складається з білків FlaA (флажелін) та FlaB, субодиниці FlaA укладені по спіралі. Порожнина усередині використовується при синтезі джгутика — він відбувається в напрямі від цитоплазматичної мембрани. Порожниною до ділянки, що збирається на даний момент, переносяться субодиниці флагеліну.
- Гачок — утворення, товстіше, ніж філамент (20-45 нм), складене з білка FlgE (можливо, також інших білків).
- Джгутиковий мотор (базальне тіло).
Джгутиковий мотор і механізм його роботи
- Основна стаття: Бактеріальний джгутиковий мотор
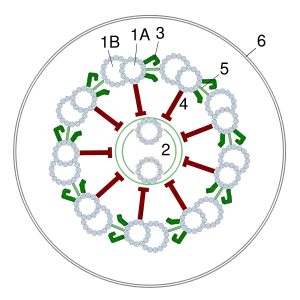
Джгутиковий мотор являє собою систему кілець, що знаходяться в цитоплазматичній та зовнішній мембранах та в клітинній стінці бактерії. Два внутрішні кільця — M (білок FliF) і S (білки FliG, FliM, FliN) (також розглядаються як єдине MS-кільце) — є обов'язковими елементами, причому M-кільце знаходиться в цитоплазматичній мембрані, а S — в периплазмі грам-негативних і пептидоглікановому шарі грам-позитивних бактерій. Ще два кільця — P і L (білки FlgH, FlgI) — присутні тільки у грам-негативних бактерій, вони розташовані в пептидоглікановому шарі і зовнішній мембрані відповідно, нерухомі і лише спрямовують стрижень ротора мотора. Навколо MS-кільця розташовані статори — білкові комплекси MotA4/MotB4 (у інших видах замість них можуть бути білки PomA, PomB, MotX та MotY), кожний з цих комплексів має протонний канал (їх може бути від 8 до 16).
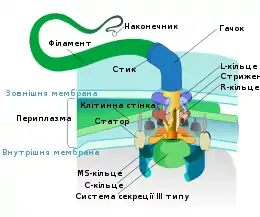
Механізм роботи джгутикового мотора дуже сильно нагадує механізм роботи трансмембранної частини АТФ-синтази або F1F0, комплексу, що синтезує АТФ та присутній у всіх живих клітинах. Існує теорія, що еволюційно джгутиковий мотор походить саме від АТФ-синтази, що скомбінувалася у процесі еволюції з системою секреції білків 3 типу. Особливістю цього мотора є можливість (у більшості видів) роботи у будь-якому напрямку та швидке перемикання між напрямками роботи.
Енергія для роботи мотора здобувається за рахунок електричного потенціалу через цитоплазматичну мембрану. Мотор пропускає протони з периплазми (або зовнішнього середовища) до цитоплазми. Деякі бактерії використовують іони натрію замість протонів (деякі морські бактерії роду Vibrio, алкалофільні Bacillus, Acetobacterium woodii). Ці іони повинні пройти через канал, розташований частково у статорі (точніше, білку FliG), а частково у роторі мотора (MotA, MotB). Через те, що частина калалу паралельна мембрані та напрямку обертання, електричний потенціал, що штовхає іон уздовж напрямку електричного поля, обертає ротор відносно статора. Наприклад, у Escherichia coli для одного оберту джгутика потрібне переміщення близько 1000 протонів. Показано, що джгутик може працювати навіть у порожніх клітинних оболонках за умови наявності електричного потенціалу на мембрані. Джгутик може обертатися зі швидкістю до 100 об/сек, при цьому напрям обертання може змінюватися менш ніж за 0,1 сек.
Механізм руху клітини
Обертання мотора викликає пасивне обертання філамента. Масивніша клітина починає обертатися приблизно із швидкістю 1/3 від швидкості джгутика у зворотному напрямі, а також набуває поступальної ходи. Переважна більшість наділених джгутиком бактерій мають паличкоподібну форму. З гідродинамічних розрахунків виходить, що для найефективнішого руху відношення довжини клітки до ширини повинне складати 3,7. Рух коків украй неефективний, тому вони частіше нерухомі.
Деякі бактерії, наприклад, спірохети, мають джгутики, розташовані в периплазмі бактерій (так звані подовжні філаменти), рух джгутиків змушує всю бактерію змінювати свою форму, за рахунок чого вона й рухається.
Швидкості руху бактерій варіюють від 20 μм/с у деяких Bacillus до 200 μм/с у Vibrio.
Регуляція руху
У ряду бактерій мотор і джгутик можуть обертатися тільки в одному напрямі, переорієнтація напрямку руху бактерії відбувається при зупинці за рахунок броунівського руху.
Для більшості бактерій важлива внутріщня асиметрія джгутиків та можливість мотора обертатися у двох напрямках. Бактерії-перитрихи збирають при русі всі свої джгутики (кожен з яких обертається проти годинникової стрілки) в один пучок. Мотори більшу частину часу обертаються у одному напрямку, але час від часу один з моторів на короткий час змінює напрямок обертання, цього звичайно достатньо, щоб розпався увесь пучок. У цей момент починається так завний «танець» бактерії, коли вона хаотично дригається на місці. Після цього, коли всі мотори знову починають обертатися в одному напрямку, джгутики знову збираються в пучок і бактерія рухається в новому напрямку.
При полярному розташуванні джгутиків один з них може штовхати бактерію, а інший тянути і навпаки, залежно від напряму обертання.
Джгутики архей
Джгутики архей відносно досить подібні до бактеріальних джгутиків. У 1980-х роках вони навіть вважалися гомологічними на підставі подібної морфології і поведінки (Cavalier-Smith, 1987). Як і у бактерій, джгутики архей складаються з філаментів, що тягнуться за межами клітини та обертаються, рухаючи клітину. Проте дослідження 1990-х років виявили численні детальні відмінності між джгутиками архей і бактерій, наприклад:
- Бактеріальні джгутики працюють за рахунок пропускання до цитоплазми протонів H+ (інколи іонів натрію Na+), архейні джгутики майже напевно витрачають енергію АТФ. Хоча мотор архей все ще не досліджений.
- У той час, коли бактеріальні клітини часто мають багато джгутиків, кожен з яких обертається незалежно, архейні джгутики складаються з пучка багатьох філаментів, які обертаються як єдине ціле.
- Бактеріальні джгутики ростуть поповненням субодиниць флажеліну на наконечнику, архейні джгутики ростуть добудовуванням судодиниць до основи.
- Бактеріальні джгутики товстіші від архейних, і бактеріальний філамінт має достатньо широкий простір усередині, через який субодиниці постачаються для росту джгутика, архейні джгутики занадто тонкі, щоб дозволити це.
- Гени багатьох компонентів бактеріальних джгутиків мають деяку схожість послідовності ДНК до компонентів системи секреції 3 типу, але компоненти бактеріальних і архейних джгутиків не поділяють ніякої схожості у послідовності. Натомість, деякі компоненти архейних джгутиків розділяють послідовність і морфологічну схожість з компонентами ворсинок 4 типу, які збираються за допомогою системи секреції 2 типу (номенклатура ворсинок/пілі та систем секреції білків не послідовна).
Ці відмінності означають, що бактеріальні і архейні джгутики — класичний випадок конвергентної еволюції, а не гомології. Проте, в порівнянні до десятиліть детального вивчення бактеріальних джгутиків (наприклад Говардом Бергом), архейні джгутики тільки недавно почали отримувати серйозну наукову увагу. Тому багато хто все ще помилково вважає, що існує тільки один основний вид джгутиків прокаріот, і що архейні джгутики належать до нього.
Джгутики еукаріотів
- Див також: ундуліподіум
Джгутики еукаріотів — абсолютно інша структура, ніж джгутики бактерій та архей, і мають зовсім інше еволюційне походження. Єдина загальна характеристика між бактеріальними, архейними та еукаріотичними джгутиками — їх зовнішній вигляд, те, що вони всі — позаклітинні структури, що використовуються для руху. Разом з війками вони складають групу органел, відомих як ундуліподії.
Джгутик еукаріотів являє собою пучок з дев'яти зв'язаних пар мікротрубочок, що оточують дві центральні мікротрубочки. Так звана структура «9+2» — характеристика ядра джгутика еукаріотів, аксонеми (або поздовжнього філаманту). В основі еукаріотичного джгутика знаходиться «базальне тіло» (також відоме як «блефаропласт» або «кінетосома»), яке є організаційним центром мікротрубочок та становить близько 500 нм у довжину. Базальне тіло за структурою ідентичне центріолі. Джгутик еукаріотів починається в межах цитоплазматичної мембрани клітини, таким чином, що внутрішній канал джгутика доступний для цитоплізми. Кожна з зовнішніх 9 пар мікротрубочок має пару дінеїнових рукояток («внутрішню» і «зовнішню») до сусідньої мікротрубочки, ці рукоятки відповідають за рух джгутика, оскільки сила, що виробляється ними, примушує пари мікротрубочок ковзати один проти одного, і джгутик згинається як одне ціле. Ці дінеїнові рукоятки отримують енергію за рахунок гідролізу АТФ. Аксонема джгутика також містить «радіальні спиці», поліпептидні комплекси, що тягнуться від кожної з зовнішніх 9 пар мікротрубочок у напрямку до центральної пари, з «головою» спиці, напрямленою всередину. Вважається, що радіальні спиці беруть участь в регулюванні руху джгутика, хоча їх точна функція і метод дії ще не відомі.
Рухомі джгутики служать для руху окремих клітин (наприклад, плаваючих протозоїв і сперматозоїдів) і транспорту рідин (наприклад, транспорт слизу стаціонарними клітинами в трахеї). На додаток до цього, нерухомі джгутики — життєво важливі органели в органах чуття та передачі сигналів у великій кількості типів клітин (наприклад, фоторецепторні палички ока, нюхові нейроні носа, кіносіліум у равлику вуха).
Внутрішньоджгутиковий транспорт (ВДТ, IFT), процес, при якому субодиниці аксонеми, трансмембранні рецептори та інші білки просуваються уздовж джгутика, істотний для належного функціонування джгутика, як руху, так і передачі сигналів.
Посилання
- Еволюція джгутиків.
- Ворсинки (включаючи фімбрії)
- Війки
- Мікротрубочка
- Мастигонема
Ресурси Інтернет
- Evolution of the Bacterial Flagella
- Molecular MachinesIndex of Illustrations, Graphics, and Animations
- Physics Today introduction to the bacterial flagellum by Howard Berg
