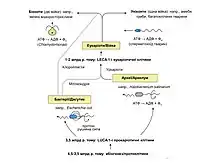
Еволюція джгутиків
Еволюція джгутиків архей, бактерій та еукаріотів є яскравим прикладом конвергентної еволюції. Вони функціонально схожі, однак, різні за походженням та розвинулися незалежно. Для позначення органели руху архей використано термін «архелум», бактерій — «джгутик», а еукаріотів — «ундуліподія».
Архелум
Джгутик архей, або архелум (отримав назву 2012 року)[2], є аналогічною структурою до бактеріального. Як і джгутик бактерій, він складається з філаменту, який, обертаючись, обумовлює рух клітини, та базального моторного комплексу, що генерує крутний момент. Хемосенсорний апарат архелума і джгутика сформований гомологічними молекулами. Проте, субодиниці філаменту архелума не демонструють гомології послідовностей із бактеріальними флагелінами, й натомість походять від білків пілей (фімбрій) IV типу. Крім того, діаметр архелума (~10 нм) ближчий до діаметра пілі (~6 нм), ніж джгутика (~25 нм).[1]
Деякі пілі IV типу здатні втягуватися, що є рушійною силою для окремих форм бактеріальної рухливості (так зване «посмикування» та «ковзання»), які дозволяють прокаріотичним клітинам пересуватися по поверхням. Втягування і витягування піля обумовлюється обертальною роботою гексамерної АТФази. В архелумі ця обертальна здатність була пристосована для здійснення плавальних рухів. Подовження філаменту архелума відбувається шляхом додавання субодиниць (пілінів) біля основи органели за участю транспортної системи ІІ типу.[3][4]
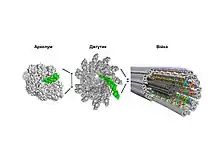
Бактеріальний джгутик
Компоненти джгутика споріднені з білками бактеріальних систем секреції. Зокрема, флагелін і джгутикові філаменти гомологічні білку EspA системи секреції III типу та EspA філаменту відповідно.[5] Всі відомі на 2012 рік неджгутикові транспортні системи III типу використовуються бактеріями для експорту (введення) токсинів у клітини еукаріотів через голковий комплекс. Схожим чином відбувається ріст джгутика, коли флагеліни експортуються через порожнистий канал органели до її кінчика, де відбувається збірка філаменту. Однак за даними філогенетичних досліджень припускають, що безпосереднім предком джгутика була більш проста система бактеріальної секреції[6].
Наразі найбільш прийнятою вважається така послідовність подій еволюційного розвитку джгутика:
- розвиток секреторної системи навколо стародавньої АТФази. Вона є спільним предком системи секреції III типу та джгутикової системи
- включення в цю структуру іонного насосу, який покращив секрецію, а згодом став моторним білком
- виникнення протофлагелярного філаменту як частини комплексу білкової секреції
- поява моторики «ковзання», яка потім вдосконалилася до плавальної моторики.[7]
Природний добір визначив еволюційну траєкторію джгутика у бік розвитку рухової функції, однак жоден з його компонентів не надає клітині селективної переваги у моториці. Даний феномен може бути пояснений за допомогою концепції експансії, яка стверджує, що сучасна функція біологічної системи відрізняється від тієї, яка існувала на стадії розвитку адаптації. Згідно з цією концепцією в процесі еволюції джгутика відбувся ряд незначних, але вирішальних функціональних зрушень, останнім з яких, імовірно, був перехід від органели адгезії або спрямованої секреції, наприклад, EspA філаменту, до вигнутої структури, здатної генерувати штовхальну силу.[5]
Ундуліподія еукаріотів
Ундуліподія — це клітинний виріст, який приблизно у 10 разів довший та складніший, ніж бактеріальний джгутик. Вона складаються з аксонеми та базального тіла, й оточена цитозольною матрицею і циліарною мембраною. Ундуліподія залучена до низки фізіологічних процесів: # рухливість клітини
- сенсорна рецепція (наприклад, сенсорні війки нейронів Caenorhabditis elegans або фоторецепторів сітківки хребетних);
- сигнало-залежний контроль внутрішньоклітинних процесів (сигналізація, опосередкована ундуліподіями, контролює деградацією та/або рециклізацію клітинних компонентів під час автофагії);
- ембріональний розвиток (формування ліво-правої асиметрії під час розвитку хребетних залежить від асиметричного потоку рідини, що рухається при обертанні нодальних війок)
- міжклітинна передача та рецепція сигналів через біоактивні позаклітинні везикули, які секретуються на кінчиках ундуліподій.
Вважають, що останній спільний предок еукаріотів (LECA), який виник 1 або 2 мільярди років тому, вже мав складну ундуліподію із рухливою аксонемою «9+2», що складалася з дев'яти зовнішніх дуплетів мікротрубочок і центральної пари синглетних мікротрубочок із асоційованими динеїновими ручками, радіальними спицями та нексиновими містками. В той час як рухливість, опосередкована роботою архелума та бактеріального джгутика, залежать від обертання філаменту під дією сили, згенереваної в основі органели, плавання за участю ундуліподій відбувається внаслідок згинання аксонеми «9+2» силами молекулярних машин, рівномірно розподілених по всій її довжині.[1] Існує три гіпотези походження ундуліподії у LECA: ендосимбіотична, вірусна та автогенна.[8]
Гіпотези ендосимбіотичного і вірусного походження
Ендосимбіотична гіпотеза (запропонована Лінн Маргуліс) стверджує, що ундуліподії походять від бактерій Gracilicutes (нащадки спірохет і простекобактерій), які вступили у постійний симбіоз із археями. Незважаючи на певні спроби популяризації гіпотези, вона не має широкої підтримки у науковому світі. Головним аргументом на її користь вважають існування еукаріотів, які використовують симбіотичні спірохети як органели руху (деякі парабазалії всередині кишечника термітів, зокрема Mixotricha і Trichonympha).[9] Водночас гомологія тубуліну мікротрубочок із бактеріальним цитоскелетним білком FtsZ, залученого в процеси реплікації, стала аргументом проти ендосимбіотичної гіпотези Магруліс, так як FtsZ-подібний білок був знайдений в клітинах архей. Це наводить на думку про ендогенне походження тубуліну.
.jpg.webp)
У 2007 році мікробіолог Пітер Сатир запропонував, що можливим попередником центріолі/базального тіла був РНК-вірус, який мав капсид із симетрією 9-го порядку та прекурсори тубуліну й тектину в своєму складі. Після зараження еукаріотичної клітини капсид вірусу перетворився на центріоль із можливістю подовження мікротрубочок.[10] На сьогоднішній день ця гіпотеза, як і попередня, не є загальноприйнятою, адже не підкріплена переконливими морфологічними та молекулярними доказами.
Гіпотеза автогенного походження
Найбільш визнана автогенна гіпотеза, висунута Джеремі Пікет-Хіпсом у 1974 році, висловлює ідею, що ундуліподія розвинулася із вже існуючих компонентів еукаріотичного цитоскелету (який має тубулін і динеїн) як надбудова апарату мітотичного веретена. Симетрія 9-го порядку цих структур могла бути результатом еволюційної оптимізації рухливості ундуліподії. Хоча віддалені гомологи тубуліну існують у прокаріотів, останні дані свідчать, що більшість компонентів ундуліподії походять від еукаріотів: 1) не було виявлено бактеріальних, архейних або вірусних аналогів центріолі/базального тіла; 2) циліарні тубуліни, мотори мікротрубочок та машинерія інтрафлагелярного транспорту є дуплікатами еукаріотичних цитоплазматичних білків.[8]
В ході біологічної еволюції відбувалася еволюція основних білкових компонентів ундуліподії.
αβ-субодиниці тубуліну, з яких утворюються мікротрубочки, мають спільного предка з прокаріотичним тубуліном FtsZ, присутнього як в клітинах архей, так і бактерій. Додаткова диверсифікація тубуліну в процесі філогенезу призвела до утворення δ-, ε- та z-ізоформ, пов'язаних із базальним тілом ундуліподії.[1]
Важкі ланцюги динеїнів належать до надродини ААА+ АТФаз і не є гомологічними до двох інших родин еукаріотичних моторних білків, кінезинів і міозинів, які мають спільного предка. Як вважається, LECA кодував різноманітний набір генів динеїну. Найімовірніше, він містив динеїни дев'яти різних підгруп: цитоплазматичний динеїн, динеїн інтрафлагелярного транспорту і сім аксонемальних динеїнів. За першим сценарієм найдавнішим динеїном був цитоплазматичний, який розвинувся в інтрафлагелярний та всі аксонемальні динеїни шляхом дуплікації генів. За другим сценарієм первинний динеїн перетворився на предка семи аксонемальних динеїнів та прототипу цитоплазматичної форми, що згодом розщепився на цитоплазматичний динеїн DHC1 і динеїн інтрафлагелярного транспорту DHC2. У першому сценарії рухлива ундуліподія могла би розвинутися з нерухомої ундуліподії із сенсорними функціями, тоді як у другому сценарії спочатку було створено рудиментарну рухливу ундуліподію, до якої на подальшому етапі були додані мотори інтрафлагелярного транспорту та сенсорні функції.[11] Перший сценарій розвитку циліарних динеїнів видається більш імовірним.
Мультимерні ІФТ (інтрафлагелярний транспорт)-асоційовані білки і BBS (Bardet–Biedl syndrome) протеїни, які транспортують аксонемальні та мембранні вантажі вздовж ундуліподії, як пропонується, мають спільне еволюційне походження із білками COPI/II, що беруть участь у внутрішньоклітинному транспорті везикул.[1]
Стародавня ундуліподія виконувала сенсорну та рухову функції, при чому першою розвинулася саме сенсорна функція. Еволюція сенсорної ундуліподії вимагала спрямованого транспорту рецепторів і сигнальних молекул до спеціалізованого мембранного домену. Тому першим етапом її розвитку вважають виникнення поляризованого везикулярного транспорту від апарату Гольджі до плазматичної мембрани за допомогою специфічних облямівок, в результаті чого утворилася сенсорна ділянка. Раннє формування дифузійного бар'єру, можливо, за участю септинів, призвело до відокремлення цієї ділянки від цитоплазми та решти плазмалеми й перетворення її на циліарну мембрану. Поляризований везикулярний транспорт, що спирався на систему моторів і адапторів, згодом перетворився в систему інтрафлагелярного транспорту.
Сенсорна ундуліподія могла використовуватися одноклітинними організмами для прикріплення до поверхонь і витягування тіла клітини в процесі т. зв. «ковзання» (gliding), що дозволяло клітинам рухатися майже амебоїдним способом. Декілька раундів дуплікації динеїну з подальшою підфункціоналізацією обумовили виникнення зовнішньої і внутрішньої ручок динеїну, які створюють силу биття джгутика. Поєднання сенсорної і рухової функцій у стародавній ундуліподії призвело до розвитку сучасної органели.[8]
Порівняння архелума, джгутика та ундуліподії
| Архелум | Джгутик | Ундуліподія | ||
|---|---|---|---|---|
| Загальна характеритика та функції | Основна функція | Просування через рідке середовище | Просування через рідке середовище | Просування через рідке середовище |
| Додаткові функції | Взаємодії клітина-клітина, прикріплення до поверхонь | Компоненти матриці біоплівки, механочутливість, пілі секреції, прикріплення до поверхонь, ковзання і роїння | Переміщення рідини, ковзання, сенсорна функція, прикріплення до поверхонь | |
| Домен життя | Археї | Бактерії | Еукаріоти | |
| Субклітинна локалізація | Мотор вбудований всередину клітини, філамент — позаклітинний | Мотор вбудований всередину клітини, філамент — позаклітинний | Подовження клітини на периферії із мікротрубочками в основі | |
| Механізм руху | Обертання спірального філаменту | Обертання спірального філаменту | Нерівномірне биття гнучкого відростку (аксонеми) | |
| Структура | Розміри | Діаметр мотору ~30 нм; діаметр суцільного філаменту ~10 нм; довжина до ~10 мкм | Діаметр мотору ~50 нм; діаметр порожнистого філаменту ~25 нм; довжина до ~10 мкм | Діаметр базального тіла ~400 нм; діаметр аксонеми ~250 нм; загальна довжина до ~100 мкм |
| Кількість білкових компонентів | ~10 | ~50 | ~500 | |
| Збирання | Родина білків збирання | ААА+ АТФази | Секреторна система бактерій ІІІ типу | Динеїни (ретроградний транспорт) і кінезини (антероградний транспорт) |
| Місце збирання | Основа | Кінчик | Кінчик | |
| Джерело енергії | Гідроліз АТФ | Протон-рушійна сила внаслідок гідролізу АТФ | Гідроліз АТФ | |
| Спосіб збирання | Приєднання пілінів в основі філаменту за невідомим механізмом | Експорт компонентів джгутика через порожнистий канал осьової структури та їхнє збирання на кінчику | Транспортування компонентів до кінчика ундуліподії кінезинами інтрафлагелярного транспорту, та зворотнє переміщення за участю динеїнів | |
| Генерування сили | Родина моторних білків | ААА+ АТФази | ExbBD/TolQR/AglQS | Динеїни |
| Місце розташування моторних білків | Основа філаменту | Основа філаменту | По всій довжині аксонеми | |
| Джерело енергії | Гідроліз АТФ | Протон-рушійна сила | Гідроліз АТФ | |
| Спосіб роботи моторних білків | Обертовий | Покроковий??? | Покроковий | |
| Гомологи та еволюція | Гомологи | Родина білків TFF (пілі IVa/b типів, система секреції ІІ типу, байндосоми, UPS пілі) | Ін'єктосоми, АТФ-синтаза, ExbBD/TolQR, MgtE, багато внутрішніх паралогічних дуплікацій | Динеїни, кінезини, тубуліни |
| Можливий шлях еволюційного розвитку | Система секреції → необертальний адгезивний піль → обертальний архелум | Система секреції білків → необертальний секреторний піль → обертальний джгутик | Транспортна система сенсорної ділянки → сенсорна органела у вигляді випинання → гнучка сенсорна органела | |
| Молекулярні механізми, що лежать в основі еволюції | Нові компоненти утворюються внаслідок дуплікації, кооптації або de novo; екзаптація наявних структур для отримання нових переваг при доборі | Нові компоненти утворюються внаслідок дуплікації, кооптації або de novo; екзаптація наявних структур для отримання нових переваг при доборі | Нові компоненти утворюються внаслідок дуплікації, кооптації або de novo; екзаптація наявних структур для отримання нових переваг при доборі |
Див. також
- Останній універсальний спільний предок
- Останній спільний предок еукаріотів
Посилання
- Khan, Shahid; Scholey, Jonathan M. Assembly, Functions and Evolution of Archaella, Flagella and Cilia. Current Biology (англ.). doi:10.1016/j.cub.2018.01.085.
- Jarrell, Ken F.; Albers, Sonja-Verena (2012-07). The archaellum: an old motility structure with a new name. Trends in Microbiology (англ.) 20 (7). с. 307–312. doi:10.1016/j.tim.2012.04.007. Процитовано 14 травня 2020.
- Faguy, David M.; Jarrell, Ken F.; Kuzio, John; Kalmokoff, Martin L. (1 січня 1994). Molecular analysis of archaeal flagellins: similarity to the type IV pilin – transport superfamily widespread in bacteria. Canadian Journal of Microbiology (англ.) 40 (1). с. 67–71. ISSN 0008-4166. doi:10.1139/m94-011. Процитовано 23 квітня 2020.
- Miyata, Makoto; Robinson, Robert C.; Uyeda, Taro Q. P.; Fukumori, Yoshihiro; Fukushima, Shun‐ichi; Haruta, Shin; Homma, Michio; Inaba, Kazuo та ін. (2020-01). Tree of motility – A proposed history of motility systems in the tree of life. Genes to Cells (англ.) 25 (1). с. 6–21. ISSN 1356-9597. PMC PMC7004002. PMID 31957229. doi:10.1111/gtc.12737. Процитовано 23 квітня 2020.
- Pallen, Mark J.; Matzke, Nicholas J. (2006-10). From The Origin of Species to the origin of bacterial flagella. Nature Reviews Microbiology (англ.) 4 (10). с. 784–790. ISSN 1740-1526. doi:10.1038/nrmicro1493. Процитовано 25 квітня 2020.
- Abby, Sophie S.; Rocha, Eduardo P. C. (27 вер. 2012). The Non-Flagellar Type III Secretion System Evolved from the Bacterial Flagellum and Diversified into Host-Cell Adapted Systems. PLOS Genetics (англ.) 8 (9). с. e1002983. ISSN 1553-7404. PMC PMC3459982. PMID 23028376. doi:10.1371/journal.pgen.1002983. Процитовано 25 квітня 2020.
- Young, Matt; Edis, Taner (2004). Why intelligent design fails: a scientific critique of the new creationism. New Brunswick, N.J.: Rutgers University Press. ISBN 0-8135-3748-7. OCLC 59717533.
- Carvalho-Santos, Zita; Azimzadeh, Juliette; Pereira-Leal, José. B.; Bettencourt-Dias, Mónica (25 липня 2011). Tracing the origins of centrioles, cilia, and flagella. The Journal of Cell Biology (англ.) 194 (2). с. 165–175. ISSN 1540-8140. PMC PMC3144413. PMID 21788366. doi:10.1083/jcb.201011152. Процитовано 25 квітня 2020.
- Margulis, Lynn, 1938-2011. (1998). Symbiotic planet : a new look at evolution (вид. 1st ed). New York: Basic Books. ISBN 0-465-07271-2. OCLC 39700477.
- Satir, Peter; Guerra, Charles; Bell, Aaron J. (2007-12). Evolution and persistence of the cilium. Cell Motility and the Cytoskeleton (англ.) 64 (12). с. 906–913. doi:10.1002/cm.20238. Процитовано 25 квітня 2020.
- Kollmar, Martin (2016-12). Fine-Tuning Motile Cilia and Flagella: Evolution of the Dynein Motor Proteins from Plants to Humans at High Resolution. Molecular Biology and Evolution (англ.) 33 (12). с. 3249–3267. ISSN 0737-4038. PMC PMC5100056. PMID 27880711. doi:10.1093/molbev/msw213. Процитовано 26 квітня 2020.
- Beeby, Morgan; Ferreira, Josie L.; Tripp, Patrick; Albers, Sonja-Verena; Mitchell, David R. Propulsive nanomachines: the convergent evolution of archaella, flagella, and cilia. FEMS Microbiology Reviews (англ.). doi:10.1093/femsre/fuaa006. Процитовано 23 квітня 2020.