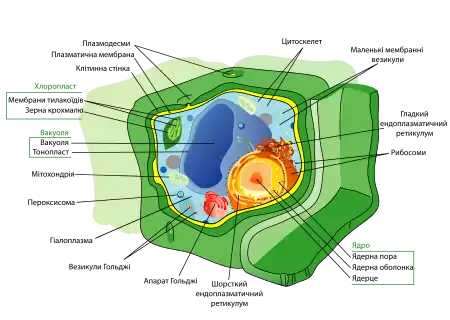
Клітинна стінка
Клітинна стінка — досить твердий шар, що оточує клітину. Вона розташовується за межами клітинної мембрани та забезпечує додаткову підтримку і захист. Клітинні стінки знайдені у бактерій, архей, грибів і рослин.
Якщо зруйнувати клітинну стінку за допомогою ферментів, залишиться протопласт — клітина, оточена клітинною мембраною. На відміну від мембран, клітинна стінка не в змозі контролювати рух молекул до клітини із зовнішнього середовища.
Клітинні стінки вищих рослин
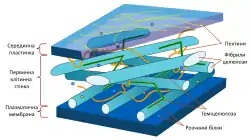
Рослинні клітинні стінки виконують цілий ряд функцій: вони забезпечують сталу форму та механічну підтримку клітини, визначаючи кінець кінцем морфологію всієї рослини. Клітинна стінка також протидіє тургору, тобто осмотичному тиску, коли до рослини поступає надлишкова кількість води. Клітинні стінки захищають від патогенів, що проникають з навколишнього середовища, та запасають для рослини вуглеводи. Рослинні клітинні стінки будуються перш за все з вуглеводного полімеру — целюлози.
Протягом цитокінезу спочатку з клітинної пластинки формується проміжна ламела, і первинна клітинна стінка розширюється усередині проміжної ламели. Фактична структура клітинної стінки не завжди чітко визначена, існують кілька моделей її структури: модель перехресних ковалентних зв'язків, модель прив'язі, модель дифузного шару і модель стратифікованого шару. Проте первинна клітинна стінка може бути визначена як мережа волокон целюлози, направлених у випадкових напрямках. Волокна утримаються разом водневими зв'язками, що забезпечує досить високу міцність. Клітини тримаються разом і розділяють желатинову мембрану, яка називається проміжна ламела та містить пектати (солі пектинової кислоти) магнію і кальцію. Клітини взаємодіють через плазмодесми, тобто цитоплазменні канальці, що з'єднують цитоплазми клітин крізь клітинні стінки.
У деяких типах клітин деяких рослин, після того як досягається максимальний розмір або певна точка розвитку, між рослинною клітиною і первинною стінкою формується вторинна стінка. На відміну від первинної стінки, волокна вирівнюються здебільшого в одному напрямку, і з кожним додатковим шаром їх орієнтація злегка змінюється. Клітини з вторинними клітинними стінками дуже тверді. Міжклітинна взаємодія все ще можлива через отвори у вторинній клітинній стінці, що дозволяють плазмодермі проникати через обидві клітинні стінки.
Основні вуглеводи, що складають первинну клітинну стінку, — це целюлоза, геміцелюлоза і пектин. Целюлозні мікроволокна зв'язуються через хеміцелюлозні мостики, формуючи целюлозно-хеміцелюлозну мережу, яка оточена матрицею пектину. Найзагальніший тип хеміцелюлози в первинній клітинній стінці — ксилоглюкан.
Рослинні клітинні стінки також містять ряд білків, найпоширеніші включають багаті на гідропролін глікопротеїни (HRGP), також відомі як екстенсіни, арабіногалактанові протеїни (AGP), багаті на гліцин протеїни (GRP) і багаті на пролін протеїни (PRP). За винятком багатих на гліцин протеїнів, всі ці білки глікозильовані і містять гідроксипролін (Hyp). Кожен клас глікопротеїнів визначається характерною, часто повторюваною послідовність амінокіслот. Химерні протеїни містять дві або більше областей, кожна з послідовністю від іншого класу глікопротеїнів. Більшість білків клітинної стінки перехресно зв'язані з вуглеводами клітинної стінки і, можливо, мають структурні функції.
Вторинні клітинні стінки можуть містити лігнін і суберин, роблячи стінки твердими. Відносний склад вуглеводів, додаткових компонентів і білків залежить від виду рослини, типу і віку клітин.
Клітинні стінки водоростей

Подібно до вищих рослин, більшість водоростей, незалежно від походження, мають клітинні стінки[1]. Клітинні стінки водоростей містять целюлозу і багато інших глікопротеїнів. Додаткові полісахариди у клітинних стінках водоростей раніше часто використовували як одну з ознак для встановлення їх таксономії, але деякі незв'язані групи мають ті ж самі полісахариди.
- Манозилові мікроволокна зустрічаються в клітинних стінках багатьох зелених водоростей (примітивних рослин), у тому числі родів Codium, Dasycladus і Acetabularia, та деяких червоних водоростей наприклад, родів Porphyra і Bangia.
- Ксілани
- Альгінова кислота — звичайний полісахарид в клітинних стінках бурих водоростей
- Сульфоновані ролісахариди зустрічаються клітинних стінках більшості водоростей, наприклад, у червоних водоростях знаходіть агарозу, карагінан, порфіран, фурселеран і фуноран.
Інші речовини, які можуть бути знайдені у водоростевих клітинних стінках, включають спорополенін і кальцій.
Група діатомових водоростей (діатомей) синтезують клітинні стінки (також відомо як фрустули або клапани) з кремнієвої кислоти (особливо ортокремневої кислоти, H4SiO4)[прояснити]. Кислота полімеризується внутріклітинно, потім стінка витісняється назовні для захисту клітини. Такі стінки вимагають менше енергії на свій синтез, даючи змогу зберегти багато енергії[2], що, можливо, частково відповідає за швидкий ріст цих організмів[3].
Клітинні стінки грибів
Не всі види грибів мають клітинні стінки, але якщо стінка присутня, вона складається з глюкозаміну і хітину, того ж вуглеводу, що надає міцність екзоскелетам комах. Вони служать подібній меті, що і клітинні стінки рослин, надаючи грибним клітинам твердість і можливість підтримувати свою форму і запобігання осмотичному лізису. Вони також обмежують вхід молекул, потенційно отруйних для гриба, наприклад деяким фунгіцидам рослинного та штучного походження. Склад, властивості і форма грибної клітинної стінки змінюються протягом життєвого циклу і залежать від умов зростання.
Інші еукаріоти
Ооміцети, патогени рослин, схожі на гриби, також мають целюлозні клітинні стінки. До недавнього часу за цю ознаку їх відносили до грибів, структурні та молекулярні свідоцтва змусили віднести їх до гетероконтів, як і бурі водорості[4].
Клітинні стінки бактерій
Як в інших організмах, бактеріальні клітинні стінки забезпечують структурну цілісність клітинам. У прокаріотів первинна функція клітинної стінки — захистити осередок від внутрішнього тургору, викликаним високою концентрацією білків та інших молекули усередині клітини порівняно з навколишнім середовищем. Бактерійна клітинна стінка відрізняється від інших тим, що її основним компонентом є пептидоглікан, шар якого розміщується негайно за межами цитоплазматичної мембрани. Пептидоглікан відповідає за твердість стінки і придання форми клітині. Він відносно пористий і не заважає потоку розчинних молекул крізь нього. Існує два основних типи бактеріальних клітинних стінок, за цією ознакою бактерії поділяються на грам-негативні і грам-позитивні.
Грам-позитивні бактерії
Грам-позитивні бактерії характеризується присутністю дуже товстого шару пептидоглікану, який відповідає за утримання фарбника кристал-віолет протягом процедури фарбування за Грамом. Така стінка знаходиться винятково в організмах, що належать до типів Actinobacteria (або грам-позитивні організми з високим вмістом G+C) і Firmicutes (або грам-позитивні організми з низьким вмістом G+C). Бактерії в межах типу Deinococcus-Thermus також можуть позитивно фарбуватися за Грамом, але містять деякі структури клітинної стінки, типові від грам-негативних організмів. В клітинну стінку грам-позитивних бактерій вбудовані полімерні спирти, тейхоєва кислота, деякі з яких зв'язуються з ліпідами, формуючи ліпотейхоєву кислоту. Ці речовини відповідають за з'єднання пептидоглікану з цитоплазматичною мембраною. Тейхоєва кислота надає клітині негативний електричний заряд завдяки наявності фосфодіестерних зв'язків між мономерами тейхоєвої кислоти.
Грам-негативні бактерії
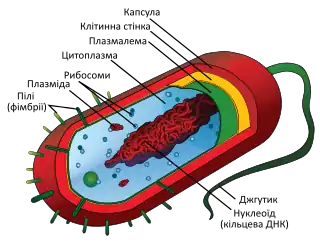
На відміну від грам-позитивних бактерій, грам-негативні бактерії містять дуже тонкий шар пептидоглікану, що відповідає за нездатність клітинних стінок утримувати фарбник кристал-віолет протягом процедури фарбування за Грамом. На додаток до шару пептидогліканів, грам-негативні бактерій мають другу, так звану зовнішню мембрану, що знаходиться назовні від клітинної стінки та компонує фосфоліпіди та ліпополісахаріди на своїй зовнішній стороні. Негативно заряджені ліпополісахаріди також надають клітині негативний електричний заряд. Хімічна структура ліпополісахарідів зовнішньої мембрани часто унікальна для окремих штамів бактерій і часто відповідає за реакцію антигенів з представниками цих штамів.
Як будь-який подвійний шар фосфоліпідів, зовнішня мембрана досить непроникна для всіх заряджених молекул. Проте білкові канали (порини), присутні в зовнішній мембрані, дозволяють пасивний транспорт багатьох іонів, цукру і амінокислот через зовнішню мембрану. Таким чином, ці молекули присутні у периплазмі — шарі між зовнішньою і цитоплазматичною мембранами. Периплазма містить шар пептидоглікану і багато білків, що відповідають за гідроліз і прийом позаклітинних сигналів. Вважається, що периплазма гелеподібна, а не рідка, через високий вміст білків і пептидоглікану. Сигнали та живильні речовини з периплазми потрапляють до цитоплазми клітини використовуючи транспортні білки в цитоплазматичній мембрані.
Клітинні стінки архей
Хоча вони і не унікальні, клітинні стінки архей дещо відрізняються від бактеріальних. Наприклад, клітинні стінки більшості архей утворені зовнішніми шарами білків або S-шаром. S-шари розповсюджені в бактеріях, де вони служать єдиним компонентом клітинної стінки в деяких організмах (наприклад в Planctomyces) або зовнішнім шаром в багатьох організмах з пептидогліканами. За винятком однієї групи метаногенів, археї не мають пептидогліканної стінки. Навіть в даному випадку пептидоглікани дуже відрізняється від типу, знайденого в бактеріях.
Примітки
- Sendbusch, P. S. (2003). Cell Walls of Algae Архівовано 28 листопада 2005 у Wayback Machine.. Botany Online
- Raven, J. A. (1983). The transport and function of silicon in plants. Biol. Rev. 58, 179—207
- Furnas, M. J. (1990). In situ growth rates of marine phytoplankton: Approaches to measurement, community and species growth rates. J. Plankton Res. 12, 1117—1151
- Interactions between Plants and Fungi: the Evolution of their Parasitic and Symbiotic Relations Архівовано 8 грудня 2006 у Wayback Machine., P. v. Sengbusch, accessed 8 December 2006