Дозова компенсація
Дозова компенсація генів — епігенетичні механізми, що дозволяють урівняти дозу зчеплених зі статтю генів у самців і самиць видів, у яких визначення статі відбувається за допомогою статевих хромосом. Так, наприклад, у самців ссавців гени X-хромосоми, не враховуючи псевдоавтосомні ділянки, наявні в одній копії, а у самиць — у двох. Оскільки така різниця могла би призвести до серйозних аномалій, існують механізми дозової компенсації генів, не пов'язаних безпосередньо із визначенням статі. У ссавців це здійснюється за допомогою інактивації однієї X-хромосоми у клітинах самиць, таким чином, що у кожній соматичній клітині особини будь-якої статі на диплоїдний набір хромосом припадає тільки одна активна X-хромосома.
Історія дослідження

1949 року Мюррей Барр і Еварт Бертрам, досліджуючи каріотип клітин котів, помітили, що у ядрах самиць наявне щільне тільце, відмінне від ядерця, проте його немає у клітинах котів чоловічої статі. Вони називали цю структуру статевим хроматином, пізніше вона стала більш відома як тільце Барра. 1959 Сусуму Оно з'ясував, що тільце Барра — це X-хромосома.
1961 Мері Лайон запропонувала гіпотезу, згідно з якою інактивація X-хромосоми забезпечує дозову компенсацію генів, а вибір хромосоми, яка буде «вимкнена», відбувається випадково. Такі висновки Лайон зробила на основі дослідження клітин із аномальною кількістю хромосом, зокрема із трисоміями по статевих хромосомах і поліплоїдією. Були отримані такі результати:
| Каріотип: | К-сть інактивованих X-хромосом |
| Нормальна жінка (XX + 44 автосоми): | 1 |
| Нормальний чоловік (XY + 44 автосоми): | 0 |
| Жінка із трисомією по X-хромосомі (XXX + 44 автосоми): | 2 |
| Чоловік із сидромом Кляйнфельтера (XXY + 44 автосоми): | 1 |
| Тетраплоїдні жіночі клітини (XXXX + 88 автосом): | 2 |

Із отриманих результатів був зроблений висновок, що клітини мають якийсь механізм «рахування» X-хромосом і залишають одну активною на пару автосом. На користь гіпотези про випадковість вибору хромосоми, що підлягатиме перетворенню у гетерохроматин, свідчило спостереження мозаїчного забарвлення гетерозиготних самиць мишей по X-зчеплених генах, що відповідають за забарвлення хутра. Мозаїчність виникає через те, що інкивація однієї із X-хромосом відбувається у клітинах зародка приблизно у період гаструляції і мітотично успадковується, таким чином клон кожної із цих клітин зберігатиме активною одну й ту ж X-хромосому: материнського або батьківського походження. Схожі результати, що свідчили про мозаїчність, були отримані при дослідженні ізоформ глюкозо-6-фосфатізомерази у людей, що кодується геном X-хромосоми.
Отримані результати швидко знайшли застосування, як у біологічних дослідженнях, так і поза їх межами. Так у 1965 Стенлі Гартлер використав інактивацію X-хромосоми, щоб довести клональне походження ракових пухлин. А 1966 року на Олімпійських іграх виявлення тілець Барра почало використовувалось для підтвердження статі спортсменів.
Найбільша кількість інформації про молекулярні механізми дозової компенсації у ссавців була накопичена завдяки роботам, проведеним на ембріональних стовбурових клітинах.
Механізми дозової компенсації
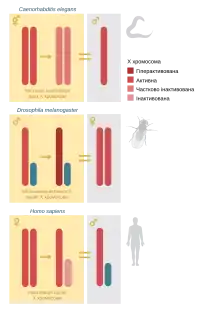
Різні види використовують різні механізми дозової компенсації: випадкову або імпринтовану інактивацію однієї із X-хромосом у гомогаметної статі (у ссавців), двократне збільшення активності експресії із однієї X-хромосоми гетерогаметної статі (у плодової мушки) або двократне її зменшення із обидвох X-хромосом гомогаметної статі (у нематоди Caenorhabditis elegans). Не залежно від конкретного механізму, компенсація завжди відбувається тільки в однієї статі і є необхідною умовою життєдіяльності особин цієї статі.
У дрозофіли і ссавців у здійснені дозової компенсації задіяні некодуючі РНК, які вкривають регульовану X-хромосому, а також здійснюється специфічна для статі модифікація білків-гістонів на відповідній хромосомі.
У птахів дозовій компенсації підлягає всього кілька генів.
У ссавців

Випадкова інактивація X-хромосоми
У плацентарних ссавців дозова компенсація генів відбувається шляхом випадкової інактивації однієї із X-хромосом. Цей процес відбувається приблизно у період гаструляції ембріонального розвитку (у людини — ~12 день). Всі нащадки певної клітини зберігають інактивованою ту ж хромосому, що й материнська клітина, тобто, як і інші епігенетичні мітки ця мітотично успадковується. Перепрограмування X-інактивації відбувається у премодріальних клітинах зародкової лінії. Ключову роль у забезпеченні інактивації X-хромосоми відіграє довга некодуюча РНК Xist, яка діє за цис-механізмом, тобто впливає на саме на ту хромосому, із якої транскрибується.
Частина генів на X-хромосомі уникають інактивації, зокрема псевдоавтосомні ділянки, тобто такі, які мають гомологічні послідовності на Y-хромосомі, і деякі інші гени, які, можливо, мають вплив на формування статевих ознак. Таких ділянок значно більше у людей, ніж у мишей. Активним на Xi залишається також ген XIST.
Оскільки через інактивацію однієї X-хромосоми доза активних X-зчеплених генів мала би бути вдвічі меншою за дозу автосомних генів, деякі дослідники вважають, що існує додатковий механізм, а саме глобальна активація всіх генів Xa (активної X-хромосоми) у 1.4—2 рази під час диференціації, хоча деякі нові дослідження із використанням високопродуктивного секвенування РНК не підтверджують її.
Імпринтована інактивація X-хромосоми
Інколи інактивація X-хромосоми може відбуватись не випадково. Наприклад, у сумчастих цьому процесу завжди підлягає хромосома батьківського походження. Це явище називається імпринтованою інактивацією і вважається еволюційно примітивнішим механізмом.
Імпринтована інактивація зрідка може виявлятись і в плацентарних ссавців, наприклад, мишей. У цих гризунів на стадії 2—4 бластомерів батьківська X-хромосома переходить у гетерохроматин. Така закономірність інактивації зберігається у трофобласті балстоцисти, який бере участь у формуванні плаценти, у той час як у клітинах ембріобласту пізніше відбувається перепрогармування: спочатку всі хромосоми переходять у активний стан, після чого відбувається випадкова інактивація.
У Drosophila melanogaster
У плодової мушки Drosophila melanogaster, хоч визначення статі, як і у ссавців, відбувається за допомогою статевих хромосом, тілець Барра у ядрах самиць немає. Натомість X-хромосома самців стає гіперактивною, рівень експресії її генів сягає сумарного рівня експресії з обидвох X-хромосом самиць. У забезпеченні такої активності ключову роль відіграє мультимерний комплекс MSL (англ. Male specific lethal), який, як свідчить його назва, є необхідним для життєдіяльності самців. До складу MSL входять білки продукти генів msl1, msl2, msl3, mle, mof, що разом із некодуючими РНК RoX1 і RoX2 формують компенсасому. Компенсасоми приєднуються до сотень ділянок єдиної X-хромосоми самців, і, ймовірно, саме вони забезпечують її гіперактивацію. Експресія принаймні деяких із msl-генів перебуває під негативним контролем продукту гену Sxl (англ. Sex lethal), що бере участь у підрахунку X-хромосом і визначенні статі.
У Caenorhabditis elegans
У нематоди C. elegans стать визначається кількістю X-хромосом: у самців вона одна, а у гермафродитів — дві. Механізм дозової компенсації у цих тварин запускається тим же сигнальним шляхом, що і визначення статі, але контролюється іншою групою генів. Полягає цей механізм у двократному зниженні активності транскрипції на обидвох X-хромосомах гермафродитів.
Визначення статі і запуск дозової компенсації починається із підрахунку кількості X-хромосом на одну автосому. X:A-сигнал у самців (X0) запускає експресію гену xol-1, що визначає розвиток зародка по чоловічому типу, а також і пригнічує експресію білка SCD-2. Наявність останнього є необхідною і достатньою умовою для активації дозової компенсації, яка, таким чином, відбувається тільки у гермафродитів. SCD-2 також забезпечує спрямування комплексу дозової компенсації до X-хромосом. Цей мультимерний комплекс містить багато білків, що мають інші функції у клітині (наприклад, MIX-1 задіяний у розходженні хромосом в анафазі), або споріднені із такими білками (зокрема, білки DPY-26, DPY-27, and DPY-28 схожі до білків конденсинового комплексу, що бере участь в упакуванні мітотичних (і мейотичних) хромосом). Комплекс дозової компенсації приєднується до X-хромосоми у кількох місцях, після чого його пригнічувальна дія може поширюватись на короткі або значні відстані.
Джерела
- Tamarin RH (2001). Principles of Genetics (вид. 7th). Mcgraw-Hill. с. 90—95. ISBN 0072334193.
- Morey C, Avner P. The demoiselle of X-inactivation: 50 years old and as trendy and mesmerising as ever // PLoS Genetics. — 2011. — July. — DOI:. — PMID:.
- Berletch JB, Yang F, Disteche CM. Escape from X inactivation in mice and humans // Genome Biology. — 2010. — Т. 11, вип. 6 (June). — DOI:. — PMID:.
- Meyer, Barbara (2005). X-Chromosome dosage compensation. WormBook (англ.). Процитовано 30 червня 2013.