Інактивація X-хромосоми
Інактивація X-хромосоми, ІXХ (англ. XIC, X-chromosome inactivation) — процес дозової компенсації генів у ссавців, який призводить до транскрипційної активності лише однієї статевої X-хромосоми у самиць та самців. Інактивація відбувається за правилом (n-1), де n — кількість X-хромосом у ядрі[1]. X-хромосома — одна з двох статевих хромосом у ссавців. У більшості ссавців самці мають статеву Y-хромосому та одну X-хромосому, тоді як у самиць — дві X-хромосоми.

Класичне визначення інактивації X-хромосоми — процес, в якому одна з двох статевих хромосом у самиць ссавців стає неактивною[2].
Однак при певних патологіях і анеуплоїдіях кількість X-хромосом може бути іншою: так при синдромі Клайнфельтера можливі варіанти істот чоловічої статі XXY, XXXY, XXXXY; при синдромі Шерешевського-Тернера самиці є моносоміками по X-хромосомі — X0, також існують самиці-трисоміки по X, — XXX. Інактивація X-хромосоми відбувається таким чином, що активною залишається лише одна X-хромосома, а всі інші перетворюються на тільця Бара. (Наприклад, у нормальної самиці XX активною буде одна X-хромосома, друга інактивується; у самця з синдромом Клайнфельтера XXXY буде активною одна X-хромосома, дві — ні).
Історія відкриття
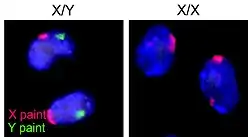
Відкриттю інактивації X-хромосоми Мері Лайон у 1961 році передувала серія відкриттів у цитогенетиці.
Роботи Теодора Бовері (англ. Theodor Heinrich Boveri) 1888 року навели сильні доводи на підтримку гіпотези, що саме хромосоми є носіями генетичної інформації в клітині. Вже 1905 року Нетті Стівенс (англ. Nettie Maria Stevens) запропонувала теорію, що статеві хромосоми відрізняються у різних статей. Едмунд Вілсон (англ. Edmund Beecher Wilson) зробив аналогічне відкриття незалежно 1905 року. 1949 року роботи Мюррея Барра (англ. Murray Llewellyn Barr) довели, що стать диференційованих соматичних клітин модельних об'єктів може бути визначено шляхом підрахування структур в ядрі, яким дали назву тільця Барра[4].
1959 року Сусумо Оно (англ. Susumu Ohno) встановив, що тільця Барра — то є X-хромосома[1]. 1959 року В. Вельшонс (англ. W. J. Welshons) і Ліане Расселл (англ. Liane B. Russell) довели, що миші-моносоміки по X-хромосомі, X0, — фенотипово нормальні, здатні до розмноження самиці, що навело на думку, що лише однієї X-хромосоми достатньо для нормального розвитку[5].
Мері Лайон (англ. Mary F. Lyon) 1961 року вивчала забарвлення хутра мишей, що є ознакою, зчепленою зі статтю, і кодується в X-хромосомі. Вона встановила, що самці XY завжди забарвлені монотонно, тоді як самиці XX можуть бути фенотиповими мозаїками — мати різнозабарвлене хутро, а самці XXY також можуть мати різне забарвлення хутра. Таким чином Мері Лайон встановила, що неактивна X-хромосома (у тільцях Барра), може бути як батьківського, так і материнського походження[1].
З нагоди 50 років відкриття інактивації X-хромосоми у липні 2011 року було проведено конференцію Європейської Організації Молекулярної Біології[6].
Механізм інактивації
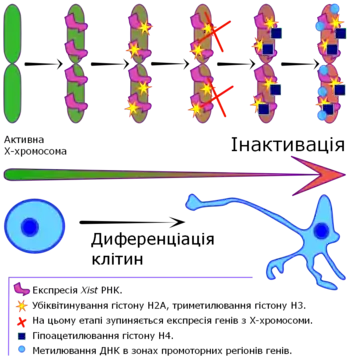
У більшості ссавців, самиці мають дві Х-хромосоми, у той час як самці мають одну Х-хромосому й одну Y-хромосому. Y-хромосома визначає стать у ранньому ембріональному періоді шляхом експресії транскрипційного фактору, що кодується SRY геном, який вмикає каскад реакцій, що призводять до фенотипу самця. За відсутності SRY розвивається фенотип самиці[8]. У самців (XY) і самиць (XX) виникає нерівновага в дозі генів, особливо враховуючи, що Y-хромосома набагато менша за Х-хромосому, і кодує лише невелику кількість генів. Інактивація X-хромосоми збалансовує таку нерівновагу.
Одна з Х-хромосом у клітинах самиць вимикається епігенетично, тобто послідовність нуклеотидів у ДНК не змінюється. Замість цього утворюється щільний гетерохроматин — фізико-хімічний стан всієї хромосоми чи її частини, в якому ускладнена взаємодія факторів транскрипції з ДНК — і процес зчитування РНК з цієї хромосоми не відбувається. Утворення гетерохроматину відбувається за допомогою метилювання ДНК та модифікації гістонових білків, а при інактивації Х-хромосоми важливу роль відіграють довгі некодуючі РНК[9].
Процес інактивації X-хромосоми складається з декількох етапів[7]:
- підрахунок X-хромосом;
- вибір хромосоми для інактивації;
- початок інактивації;
- підтримання X-хромосоми в неактивному стані.
У подальшому неактивна X-хромосома залишається стабільно заглушеною. У цьому важливу роль грає метилювання ДНК — епігенетичний процес, що полягає в додаванні метильної групи до нуклеотиду цитозину. Така біохімічна зміна може підтримуватися довгий час і впливати на активність генів[10].
Компоненти гетерохроматину неактивної X-хромосоми відрізняються від гетерохроматину в інших хромосомах. На неактивній X-хромосомі знайдено варіант гістонового білку macroH2A, та білок Trithorax. Також встановлено, що, на відміну від інших хромосом, білкові компоненти неактивної X-хромосоми розподілено рівномірно по всій її довжині[7][11].
Вивчення інактивації X-хромосоми пролило світло на декілька молекулярно-біологічних процесів: роль довгих некодуючих РНК, геномний імпринтинг та соматичне парування хромосом у ссавців[1].
Методика RAP–MS (англ. RNA antisense purification followed by quantitative mass spectrometry) дозволяє вивчати in vivo взаємодію білків та довгих некодуючих РНК. За допомогою RAP–MS у 2015 році встановили, що задля розміщення днРНК Xist на хромосомі, необхідна дія білку SAFA[12] (англ. scaffold attachment factor A). Крім цього, виключення (нокдаун) генів, що кодують білки, SHARP[13](англ. SMRT and HDAC1-associated repressor protein) та LBR[14](англ. lamin-B receptor) призводило до зупинки інактивації X-хромосоми у дослідах над ембріональними стовбуровими клітинами мишей[15][16].
При розміщенні Xist на X-хромосомі з цією хромосомою вже не з'єднується РНК-полімераза II — полімераза, яка транскрибує більшість мРНК. Виключення гену, що кодує SAFA призводило до хаотичного розміщення Xist, тоді як виключення гену, що кодує білок SHARP призводило до повернення РНК-полімерази II. Також білок SHARP взаємодіє з білками-ремодуляторами структури хроматину, такими як гістондеацетилази. Причому, виключення гістондеацетилази 3 (HDAC3), а не інших видів гістондеацетилаз, призводило до порушення механізму інактивації X-хромосоми[15][16].
Важливим елементом інактивації є дія репресивного комплексу Polycomb, PRC2 (англ. Polycomb repressive complex 2), проте дія комплексу PRC2 не важлива при ініціації процесу інактивації, скоріше у підтриманні хромосоми у інактивованому стані – триметилюванні 27 лізину H3 гістону (H3K27me3, див табличку "Порівняння еу- та гетерохроматину")[15]
ЦІX — центр інактивації X-хромосоми
Дослідження на мишачих моделях встановили, що для інактивації X-хромосоми потрібна особлива ділянка — центр інактивації X-хромосоми, ЦІX (англ. XIC, X inactivation centre)[17]. Центр інактивації X-хромосоми складається приблизно з мільйона пар основ, має декілька елементів, що беруть участь у інактивації X-хромосоми, та містить принаймні чотири гени[18]. Для початку інактивації потрібно два таких центри, по одному на кожній хромосомі, і необхідно, щоб між ними був зв'язок. Взаємодія між двома гомологічними X-хромосомами відбувається в центрі інактивації. Але залишається відкритим питання, що саме є причиною, а що наслідком: чи зближення хромосом призводить до початку інактивації, чи навпаки[19].
Xist
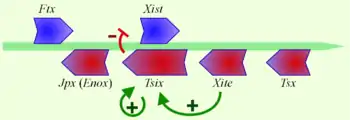
У межах ділянки центру інактивації X-хромосоми закодовано ген Xist (англ. X-inactive specific transcript), який транскриптується в довгу некодуючу РНК Xist. Xist вкриває ту X-хромосому, що буде неактивною (спочатку в зоні ЦІX, а потім — по всій довжині хромосоми). В процесі ембріонального розвитку Xist експресується з обох хромосом, але потім в одній X-хромосомі експресія Xist припиняється (і саме ця хромосома залишиться активною). Придушення експресії Xist збігається в часі з початком інактивації X-хромосоми[17].
При гетерозиготній мутації Xist, тобто такій, коли нормальний Xist є лише на одній з двох гомологічних хромосом, та X-хромосома, в якій є мутантний Xist, не інактивується[20].
Tsix
З локусу центру інактивації X-хромосоми зчитується антизмістовний транскрипт з комплементарного ланцюга ДНК того ж гену Xist. Такій нкРНК дали назву Tsix (Xist написаний задом-наперед), і встановили, що Tsix — негативний регулятор Xist, і його експресія необхідна для підтримки X-хромосоми в активному стані. Багато праць вказують на те, що саме співвідношення Tsix/Xist важливе для вибору, який алель буде глушитись, і, відповідно, яку хромосому буде інактивовано[17]. Є дані, що саме Tsix призводить до асоціації двох гомологічних X-хромосом, і експресія Tsix РНК є необхідною, але не достатньою умовою для підрахунку й вибору хромосоми для інактивації[19]
Tsix стала першою відомою антизмістовною РНК ссавців, що зустрічається в природі та має чітку функцію in vivo.
Додаткові регулятори
У зоні центру інактивації X-хромосоми знайдено велику кількість ділянок, які впливають на процес ІXХ. Такі ділянки впливають на процес інактивації X-хромосоми як у цис-, так і в транс-положенні, тобто як на одній і тій же хромосомі, на якій вони розташовані (цис-регуляторний елемент), так і на іншій (транс-регуляторний елемент). Багато некодуючих РНК впливають на активність Xist та Tsix (Jpx, Ftx та Tsx).
Xite
Xite (англ. X-inactivation intergenic transcription element) — ще один некодуючий транскрипт, що розташований перед Tsix і діє як підсилювач експресії Tsix на майбутній активній X-хромосомі[21].
LINE1
У геномі людини значну частку всієї послідовності ДНК становлять так звані транспозони, або мобільні елементи геному. Частина їх є ретротранспозонами (у людини ретротранспозони займають до 42% геному) — мобільними елементами, які копіюють і вставляють себе в геном за допомогою транскрипції з ДНК на РНК, а потім зворотної транскрипції з РНК у ДНК. LINE1 (англ. Long Interspersed Nuclear Elements) — один з активних ретротранспозонів у людини[22]. LINE1 зустрічається в X-хромосомі набагато частіше, ніж в інших хромосомах. Є праці, які вказують на участь РНК LINE1 в інактивації X-хромосоми[23].
Серія активацій та інактивацій X-хромосом
На початкових стадіях розвитку є різниця, якого походження X-хромосома, батьківського чи материнського. З початку ембріогенезу X-хромосома батьківського походження завжди неактивна. В цьому процесі важливу роль грає геномний імпринтинг. Потім, у період формування бластули обидві X-хромосоми активуються[9]. У подальшому розвитку в ембріональних клітинах інактивація X-хромосом відбувається в довільному порядку, незалежно від походження X-хромосом. Але в позаембріональних тканинах (включно з трофобластом, що формує більшу частину плаценти) активною залишається лише X-хромосома від матері, а батьківська X-хромосома інактивується[1].
Далі в зародку при формуванні майбутніх статевих клітин (гаметогенез) відбувається наступний етап активацій X-хромосом перед мейотичним поділом. Кожна з X-хромосом отримує постійну імпринтну мітку, що вказує, якого вона походження.
Гени, що зчитуються з неактивної X-хромосоми
Деякі гени, що розташовані на неактивній X-хромосомі, уникають заглушення й експресуються з обох X-хромосом. У людській лінії фібробластів 15% генів, що розташовані на неактивній X-хромосомі, тією чи іншою мірою експресуються. Рівень зчитування цих генів дуже залежить від того, в якій частині хромосоми вони закодовані[24]. Такі гени призводять до різноманіття, яке залежить від статі й типу тканини[25].
Інактивація X-хромосоми в різних видів
Основні роботи з вивчення інактивації X-хромосоми було зроблено на мишах. В останні роки надходить усе більше даних, що мишача модель інактивації X-хромосоми відрізняється від інших ссавців.
У кроликів та людей Xist-гомолог не підлягає імпринтингу, Xist зчитується з обох хромосом. У кроликів це може ввімкнути процес ІXХ на обох X-хромосомах[26].
Більше того, X-хромосоми в багатьох видів мають досить специфічний набір генів: такі гени мають низький рівень експресії в соматичних тканинах, але високий рівень експресії — у тканинах, які беруть участь у репродуктивних функціях організму (наприклад, яєчники)[25].
XACT РНК у людини
2013 року дослідники РНК людини виявили довгу некодуючу РНК XACT (англ. X-active coating transcript), що зв'язується з активною X-хромосомою[27]. XACT експресується з активної X-хромосоми, але при диференціації глушиться, і вже в диференційованих клітинах (таких, як фібробласти) XACT РНК немає. За відсутності XIST-РНК, XACT експресується з обох X-хромосом у людини, але не в миші[28].
Випадковість вибору X-хромосоми
Раніше вважалося, що вибір хромосоми для інактивації повністю випадковий, і кожна з двох гомологічних X-хромосом буде інактивована з імовірністю 50%. Але з'явилися публікації, які доводять, що в деяких модельних організмах генетичні фактори впливають на вибір. Так, у мишей існують регуляторні елементи (англ. Xce, X-controlling element), які мають три алельні форми, і одна з них, Xcec, частіше зустрічається в активній X-хромосомі, тоді як Xcea частіше зустрічається в неактивній[30].
Залишається нез'ясованим, чи в довільному порядку відбувається інактивація X-хромосоми в людини. Нещодавні дослідження вказують на те, що генетичне оточення може впливати на вибір X-хромосоми, яку буде інактивовано[30].
Див. також
Джерела
- Jeannie T. Lee (2011). Gracefully ageing at 50, X-chromosome inactivation becomes a paradigm for RNA and chromatin control. Nature Reviews Molecular Cell Biology 12: 815–826. PMID 22108600. doi:10.1038/nrm3231.
- Handbook of Epigenetics. Elsevier Inc. 2011. с. 341–351. ISBN 978-0-12-375709-8.
- Janice Y Ahn, Jeannie T Lee (2010). Retinoic acid accelerates downregulation of the Xist repressor, Oct4, and increases the likelihood of Xist activation when Tsix is deficient. BMC Developmental Biology. PMID 20727175. doi:10.1186/1471-213X-10-90.
- Murrey L. Barr & Ewart G. Bertram (Apr 1949). A morphological distinction between neurones of the male and female, and the behaviour of the nucleolar satellite during accelerated nucleoprotein synthesis. Nature 163: 676–677. PMID 18120749. doi:10.1038/163676a0.
- Mary F. Lyon (1962). Sex Chromatin and Gene Action in the Mammalian X-Chromosome. Am J Hum Genet 14 (2): 135–148. PMID 14467629.
- 50 Years of X–inactivation Research – EMBO. Oxford, United Kingdom. 20-24 July 2011. Архів оригіналу за 17 серпня 2013. Процитовано 15 травня 2013.
- Anton Wutz (2011). Gene silencing in X-chromosome inactivation: advances in understanding facultative heterochromatin formation. Nature Reviews Genetics: 542–553. PMID 21765457. doi:10.1038/nrg3035.
- Wolf Reik & Annabelle Lewis (2005). Co-evolution of X-chromosome inactivation and imprinting in mammals. Nature Reviews Genetics 6: 403–410. PMID 15818385. doi:10.1038/nrg1602.
- Wolf Reik, Anne C. Ferguson-Smith (2005). Developmental biology: The X-inactivation yo-yo. Nature 438: 297–298. PMID 16292295. doi:10.1038/438297a.
- Susan C. Wu & Yi Zhang (2010). Active DNA demethylation: many roads lead to Rome. Nature Reviews Molecular Cell Biology 11: 607–620. PMID 20683471. doi:10.1038/nrm2950.
- А. В. Сиволоб, К. С. Афанасьєва, Молекулярна організація хромосом. — К. : Видавничо-поліграфічний центр «Київський університет», 2012, 182 с. (стор. 27)
- Білок SAF-A (інші назви: Hnrnpu, Heterogeneous nuclear ribonucleoprotein U) миші в датабазі UniProt Q8VEK3
- Білок Sharp (інші назви: Spen, Msx2-interacting protein) миші в датабазі UniProt Q62504
- Білок Lbr миші в датабазі UniProt Q3U9G9
- Zlotorynski Eytan (2015). Non-coding RNA: X chromosome inactivation unravelled. Nature Reviews Genetics 16 (6): 315–315. ISSN 1471-0056. doi:10.1038/nrg3955.
- Colleen A. McHugh, Chun-Kan Chen, Amy Chow, et al (May 2015). The Xist lncRNA interacts directly with SHARP to silence transcription through HDAC3. Nature 521 (7551): 232–236. PMID 25915022. doi:10.1038/nature14443.
- Sandrine Augui, Elphège P. Nora & Edith Heard (2011). Regulation of X-chromosome inactivation by the X-inactivation centre. Nature Reviews Genetics 12: 429–442. PMID 21587299. doi:10.1038/nrg2987.
- Philip Avner & Edith Heard (2001). X-chromosome inactivation: counting, choice and initiation. Nature Reviews Genetics 2: 59–67. PMID 11253071. doi:10.1038/35047580.
- Céline Morey and Wendy Bickmore (2006). Sealed with a X. Nature Cell Biology 8: 207–209. PMID 16508671. doi:10.1038/ncb0306-207.
- Monkhorst K, Jonkers I, Rentmeester E, Grosveld F, Gribnau J. (2008). X Inactivation Counting and Choice Is a Stochastic Process: Evidence for Involvement of an X-Linked Activator. Cell 132: 410–421. PMID 18267073. doi:10.1016/j.cell.2007.12.036.
- Tim R Mercer & John S Mattick (2013). Structure and function of long noncoding RNAs in epigenetic regulation. Nature Structural & Molecular Biology 20: 300–307. PMID 23463315. doi:10.1038/nsmb.2480.
- Henry L. Levin & John V. Moran (2011). Dynamic interactions between transposable elements and their hosts. Nature Reviews Genetics 12: 615–627. PMID 21850042. doi:10.1038/nrg3030.
- Jennifer C. Chow, Constance Ciaudo, Melissa J. Fazzari, Nathan Mise, Nicolas Servant, Jacob L. Glass, Matthew Attreed, Philip Avner, Anton Wutz, Emmanuel Barillot, John M. Greally, Olivier Voinnet & Edith Heard (June 2010). LINE-1 activity in facultative heterochromatin formation during X chromosome inactivation. Cell 141 (6): 956–969. PMID 20550932. doi:10.1016/j.cell.2010.04.042.
- Laura Carrel1 & Huntington F. Willard (2005). X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 434: 400–404. PMID 15772666. doi:10.1038/nature03479.
- Deng X, Hiatt JB, Nguyen DK, Ercan S, Sturgill D, Hillier LW, Schlesinger F, Davis CA, Reinke VJ, Gingeras TR, Shendure J, Waterston RH, Oliver B, Lieb JD, Disteche CM. (2011). Evidence for compensatory upregulation of expressed X-linked genes in mammals, Caenorhabditis elegans and Drosophila melanogaster. Nature Genetics 43: 1179–1185. PMID 22019781. doi:10.1038/ng.948.
- Okamoto I, Patrat C, Thépot D, Peynot N, Fauque P, Daniel N, Diabangouaya P, Wolf JP, Renard JP, Duranthon V, Heard E. (2011). Eutherian mammals use diverse strategies to initiate X-chromosome inactivation during development. Nature 472: 370–374. PMID 21471966. doi:10.1038/nature09872.
- Inês Chen (2013). A lncRNA for the active X. Nature Structural & Molecular Biology. doi:10.1038/nsmb.2538.
- Céline Vallot, Christophe Huret, Yann Lesecque, Alissa Resch, Noufissa Oudrhiri, Annelise Bennaceur-Griscelli, Laurent Duret & Claire Rougeulle (2013). XACT, a long noncoding transcript coating the active X chromosome in human pluripotent cells. Nature Genetics 45: 239–241. PMID 23334669. doi:10.1038/ng.2530.
- Grant J, Mahadevaiah SK, Khil P, Sangrithi MN, Royo H, Duckworth J, McCarrey JR, VandeBerg JL, Renfree MB, Taylor W, Elgar G, Camerini-Otero RD, Gilchrist MJ, Turner JM. (2012). Rsx, a metatherian RNA with Xist-like properties. Nature 487: 254–258. PMID 22722828. doi:10.1038/nature11171.
- Renault NK, Pritchett SM, Howell RE, Greer WL, Sapienza C, Orstavik KH, Hamilton DC. (2013). Human X-chromosome inactivation pattern distributions fit a model of genetically influenced choice better than models of completely random choice. European Journal of Human Genetics. PMID 23652377. doi:10.1038/ejhg.2013.84.
Література
- X Inactivation and Epigenetics. YouTube (англ.). WEHImovies. 2012. Процитовано 16 липня 2013.
- Morey C, Avner P. The demoiselle of X-inactivation: 50 years old and as trendy and mesmerising as ever // PLoS Genetics. — 2011. — July. — DOI:. — PMID:. Процитовано 19.07.2013.
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр «Київський університет». с. 194.
- А. В. Сиволоб, К. С. Афанасьєва (2012). Молекулярна організація хромосом. К: Видавничо-поліграфічний центр «Київський університет». с. 27.
|
|
Ця стаття належить до добрих статей української Вікіпедії. |
