Клітинне ядро
У клітинній біології, ядро (лат. nucleus) — клітинна органела, знайдена у більшості клітин еукаріотів і містить ядерні гени, які складають більшу частину генетичного матеріалу. Ядро має дві первинні функції: керування хімічними реакціями в межах цитоплазми і збереження інформації, потрібної для поділу клітини.
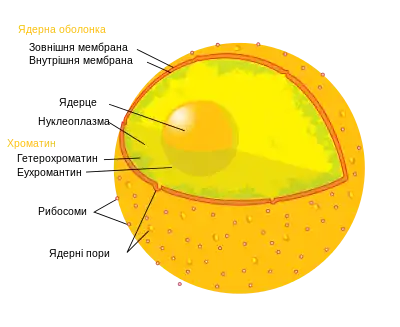
Окрім клітинного геному, ядро містить певні білки, які регулюють зчитування генетичної інформації. Зчитування гена на ядерному рівні залучає складні процеси транскрипції, обробки первинної мРНК і експорт зрілої мРНК до цитоплазми.
Ядро звичайно має розмір 8-25 мікрометрів у діаметрі. Воно оточено подвійною мембраною, яка називається ядерною оболонкою. Крізь внутрішню і зовнішню мембрани на деяких інтервалах проходять ядерні пори. Ядерна оболонка регулює і полегшує транспорт між ядром і цитоплазмою, відокремлюючи хімічні реакції, що відбуваються в цитоплазмі, від реакцій, що трапляються в межах ядра. Зовнішня мембрана безперервна з гранулярним ендоплазматичним ретикулумом (англ. RER) і може мати зв'язані рибосоми. Простір між двома мембранами (який називається «перинуклеарним простором») безперервний з люменом RER. Ядерна сторона ядерної оболонки оточена мережею проміжних філаментів, яка називається ядерною ламіною.
Внутрішня частина ядра містить одне або декілька ядерець, оточених матрицею, яка називається нуклеоплазмою. Нуклеоплазма (каріолімфа, ядерний сік, каріоплазма) — гелеподібна рідина (подібна у цьому відношенні до цитоплазми), в якій розчинені багато речовин. Ці речовини включають нуклеотид-трифосфати, сигнальні молекули, ДНК, РНК та білки (ензими та філаменти).
Генетичний матеріал присутній в ядрі у вигляді хроматину, або комплексу білка і ДНК. ДНК присутня як цілий ряд дискретних молекул, відомих як хромосоми. Є два види хроматину: еухроматин і гетерохроматин. Еухроматин — менш компактна форма ДНК. Області ДНК, які знаходяться у формі еухроматину містять гени, які часто зчитуються клітиною.
У гетерохроматині ДНК більш компактно упакована. Області ДНК, які знаходяться у формі гетерохроматину містять гени, які не зчитуються клітиною на даній стадії розвитку (цей вид гетерохроматину відомий як факультативний гетерохроматин) або є областями, які складають теломери і центромери хромосом (цей вид гетерохроматину відомий як конструктивний гетерохроматин). У багатоклітинних організмах, клітини надзвичайно спеціалізовані, щоб виконувати специфічні функції, тому різні набори генів потрібні і зчитуються. Тому, області ДНК, які знаходяться у формі гетерохроматину, залежать від типу клітини.
Ядерце — щільна структура в ядрі, де збираються елементи рибосом. У ядрі може бути одне або декілька ядерець.
Історія відкриття
В 1831 році англійський природознавець Роберт Броун вивчав різні види рослин, зразки яких він зібрав під час подорожі до Австралії. Броун був дуже уважним до деталей, а клітини рослин особливо цікавили його. Розглядаючи їх під мікроскопом, він побачив дещо цікаве: кожна клітина містила круглий і непрозорий елемент. Він назвав його ядром.
Дізнавшись про спостереження Броуна, німецький фізіолог Теодор Шванн почав шукати подібні елементи в клітинах пуголовків і знайшов. Кожна клітина містила ядро. Це був революційний прорив — свідчення того, що всі види життя пов'язані між собою. В одній із книг Шванн описав різні типи клітин, взяті від різноманітних організмів і визначив їх за фактом наявності ядра.
Усвідомлення того, що є елемент спільний для всіх організмів, не тільки для рослин, а й для тварин, поєднало рослинне і тваринне царство у щось спільне, щось, що мало однакові риси.
Тонка структура клітинного ядра
Хроматин
Величезна довжина молекул ДНК еукаріот визначила появу спеціальних механізмів зберігання, реплікації і реалізації генетичного матеріалу. Хроматином називають молекули хромосомної ДНК в комплексі зі специфічними білками, необхідними для здійснення цих процесів. Основну масу складають «білки зберігання», так звані гістони. З цих білків побудовані нуклеосоми — структури, на які намотані нитки молекул ДНК. Нуклеосоми розташовуються досить регулярно, так що утворюється структура, яка нагадує намисто. Нуклеосома складається з білків чотирьох типів: H2A, H2B, H3 та H4. В одну нуклеосому входять по два білка кожного типу — всього вісім білків. Гістон H1, більший ніж інші гістони, зв'язується з ДНК в місці її входу на нуклеосому. Нуклеосома разом з H1 називається хроматосомою.
Якщо хроматин упакований щільно, його називають конденсованим або гетерохроматином, його можна побачити під мікроскопом. ДНК, що знаходиться в гетерохроматині. У інтерфазі гетерохроматин звичайно розташовується по периферії ядра (пристінковий гетерохроматин). Повна конденсація хромосом відбувається перед поділом клітини. Якщо хроматин упакований нещільно, його називають еу- або інтерхроматином. Цей вид хроматину набагато менш щільний при спостереженні під мікроскопом і зазвичай характеризується наявністю транскрипційної активності. Щільність упаковки хроматину є частиною епігенетичного контролю експресії генів і частково визначається модифікаціями гістонових хвостів — ацетилюванням та деацетилюванням і метилюванням.
Вважається, що в ядрі існують так звані функціональні домени хроматину (ДНК одного домену містить приблизно 30 тисяч пар основ), тобто кожна ділянка хромосоми має власну «територію». На жаль, питання просторового розподілу хроматину в ядрі вивчений поки недостатньо. Відомо, що теломерні (кінцеві) і центромерні (що відповідають за зв'язування сестринських хроматид в мітозі) ділянки хромосом закріплені на білках ядерної ламіни.
Еволюційне значення клітинного ядра
Основна функціональна відмінність клітин еукаріот від клітин прокаріотів полягає в просторовому розмежуванні процесів транскрипції (синтезу матричної РНК) і трансляції (синтезу білка рибосомою), що дає в розпорядження еукаріотичної клітини нові інструменти регуляції біосинтезу і контролю якості мРНК.
У той час, як у прокаріотів мРНК починає транслюватися ще до завершення її синтезу РНК- полімеразою, мРНК еукаріотів зазнає значних модифікації (так званий процесинг), після чого експортується через ядерні пори в цитоплазму, і тільки після цього може вступити в трансляцію. Процесинг мРНК включає декілька елементів.
З попередника мРНК (пре-мРНК) в ході процесу, званого сплайсингом вирізаються інтрони — незначущі ділянки, а значущі ділянки — екзони з'єднуються один з одним. Причому екзони однієї і тієї ж пре- мРНК можуть бути з'єднані декількома різними способами (альтернативний сплайсинг), так що один попередник може перетворюватися в декілька різних зрілих мРНК. Таким чином, один ген може кодувати відразу декілька білків.
Модифікаціям піддаються кінці молекули мРНК. До 5'-кінця молекули прикріплюється 7-метилгуанін (так званий кеп). До 3'- кінця приєднуються кілька десятків залишків аденіну (поліаденілування). Також за допомогою альтернативного поліаденілування можна контролювати наступну долю мРНК, наприклад в ході РНК інтерференції — адже 5'- та 3'- нетрансльовані ділянки є місцями з'єднання мікроРНК[1].
Примітки
- Ran Elkon, Alejandro P. Ugalde & Reuven Agami (July 2013). Alternative cleavage and polyadenylation: extent, regulation and function. Nature reviews. Genetics 14 (7): 496–506. PMID 23774734. doi:10.1038/nrg3482.
