Міксоспоридії
Міксоспоридії (Myxosporea) — клас мікроскопічних паразитів, що належать до групи Myxozoa. Вони локалізуються в порожнинах внутрішніх органів і різних тканинах хазяїна; можливе й внутрішньоклітинне паразитування, наприклад, в ікрі риб. Мають складний біологічний життєвий цикл з розвитком вегетаційної форми або в двох хазяях: водних безхребетних (переважно кільчасті черви) і хребетних, зазвичай рибах, або ж лишень в одному хазяїні — рибі. Кожний хазяїн звільняє різні типи спор. Дві форми спор є настільки різними, що раніше розглядались, як такі що належать до різних класів у межах Myxozoa. Описано понад тисячу видів міксоспоридій.
? Міксоспоридії | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Цисти Henneguya zschokkei у м'ясі лосося | ||||||||||
| Біологічна класифікація | ||||||||||
| ||||||||||
Ряди[1] | ||||||||||
| ||||||||||
Посилання | ||||||||||
| ||||||||||
Життєвий цикл
Міксоспоридна стадія
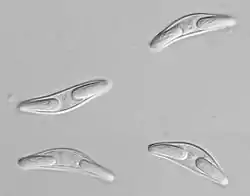
У хребетного хазяїна, міксоспоридії характеризується спорами, що складається з кількох клітин. Спори мають досить складну будову. Спори ззовні вкриті оболонкою з 2-6 стулок. Всередині містяться 1-2 амебоїдні інфекційні зародкові клітин (спороплазма) з двома гаплоїдними ядрами, а також від 2 до 7 подібних на нематоцисти полярні капсули з жалкими нитками. У деяких стадій життєвого циклу статеві клітини повністю заповнюють зміст клітин хазяїна. Найпростіші міксоспоридії мають дві стулки у вигляді напівсфер, що з'єднані по екватору швом. Стулки можуть бути сплющеними та витягнутими або утворювати на полюсах довгі тонкі вирости. Однак переважно спори мають веретеноподібну, лінзоподібну або сигароподібну форми. Іноді спора складається з трьох — шести стулок і набуває зірчастої форми. Оболонка спори складається з оболонок капсул, які з'єднані між собою вздовж лінії шва. Деякі види містять запаси полісахаридів у вигляді частинок β-глікогену, які концентруються в центральній «йодофілній вакуолі».
Стулки можуть мати гладеньку або ребристу поверхню, або бути витягнуті у латеральній проекції, можлива наявність слизової оболонки. Ці адаптація, ймовірно, служать для збільшення плавучості спори в товщі води. Клапани формуються із стійких некератинизованих білків.

Полярні, або жалкі, капсули — це округлі, грушоподібні або овальні утвори. Найчастіше в спорі містяться дві такі капсули, хоча в деяких видів кількість їх відрізняється. Усю порожнину капсули займає вакуоля, всередині якої розташована спірально скручена полярна, або жалка, нитка; один кінець її вільний, другий прикріплений до краю капсули. На передньому звуженому кінці капсули є отвір, що прикривається невеличкою кришечкою, в центрі якої є невелика ділянка — корочок, що, ймовірно, відрізняється від кришечки за хімічним складом. Кришечки полярних капсул видаються назовні спори через спеціальні отвори в стінках стулок поблизу лінії швів. Завдяки такій будові кришечка під час надходження до травної системи безпосередньо зазнає дії травних ферментів хазяїна, які руйнують корочок. Це ймовірно й призводить до викидання (вистрілювання) жалкої нитки, однак механізм цього процесу повністю не з'ясований. На відміну від мікроспоридій полярні жалкі нитки міксоспоридій призначені не для транспорту амебоїдного зародка, а для утримання спори на стінці кишечника.
Спори міксоспоридій дуже стійкі. Відомо, що спори Myxosoma cerebralis, що паразитують у форелі та інших лососевих рибах, протягом 12 років залишались живими на дні висохлої водойми. Стулки спор добре витримують також дію різних хімічних речовин. Зруйнувати їх вдається лише шляхом кип'ятіння в сірчаній кислоті. Після прикріплення спори до стінки кишечника її, стулки розкриваються під дією травних ферментів хазяїна, амебоїдний зародок виходить у просвіт кишечника, а звідти потрапляє до місця паразитування. В амебоїдному зародку, що перебуває в спорі або мігрує в організмі хазяїна, гаплоїдні ядра зливаються в одне диплоїдне. Цей процес називається автогамією й веде до відновлення диплоїдності. Імовірно, що у предків міксоспоридій відбувався справжній статевий процес і перехресний обмін хромосомами, однак у процесі історичного розвитку внаслідок паразитичного способу життя, що утруднює зустріч гамет, копуляція звелася до відновлення числа хромосом.
Життєвий цикл міксоспоридій проходить з гаметичною редукцією. У місці локалізації диплоїдний амебоїдний зародок росте. Його ядро багаторазово мітотично ділиться, внаслідок чого утворюється багатоядерний плазмодій. Уже під час перших поділів його ядра диференціюються на вегетативні (соматичні) та генеративні. Вегетативні ядра згодом стають поліплоїдними, генеративні залишаються диплоїдними. Крупніші вегетативні ядра утворюють енергіди та беруть участь у синтезі білків, менші — генеративні ядра— енергід не утворюють і в обміні речовин участі не беруть. Отже, у міксоспоридій спостерігається ядерний дуалізм. Вважалося, що генеративні ядра, як і вегетативні, містяться в цитоплазмі плазмодія, однак результати електронно-мікроскопічних досліджень свідчать, що навколо них є цитоплазма, оточена плазмалемою. Таким чином, у багатоядерному плазмодії, що виконує роль вегетативного тіла (соми), розташовані генеративні клітини, тобто, подібно до вольвокса, плазмодій — багатоклітинний організм. Плазмодій рухається за допомогою лобоподій, живиться шляхом піноцитозу або утворює численні тоненькі відростки — мікроворсинки, через плазмалему яких назовні виділяються ферменти: прилеглі тканини хазяїна перетравлюються та всмоктуються мікроворсинками (пристінкове травлення). У деяких видів спостерігається фагоцитоз клітин хазяїв.
Плазмодії розмножуються нестатево шляхом поділу та зовнішнього й внутрішнього брунькування. Перед спороутворенням генеративні клітини (одна або кілька) оточуються ще однією мембраною, утворюючи панспоробласт. У клітинах панспоробласту ядро послідовно мітотично ділиться на вісім диплоїдних ядер, два припиняють поділ і перетворюються на соматичні ядра, які керують вегетативними функціями. Шість інших ядер діляться ще раз шляхом мейозу (одноступеневий мейоз), утворюючи 12 гаплоїдних, що групуються по шість у споробласти, які відрізняються від панспоробластів тим, що не мають соматичних ядер. У споробластах навколо ядер локалізуються ділянки цитоплазми, і вони, стають багатоклінними.
Типи нестатевого розмноження мікроспоридій: поділ багатоядерного плазмодія, зовнішнє та внутрішнє брунькування, утворення панспоробластів, перетворення амебоїдного зародка в багатоядерний плазмодій: вегетативне ядро; генеративна клітина з генеративним ядром; споробласт тинними. Із двох клітин формуються стулки, після чого ядра руйнуються, дві клітини переміщуються до одного з полюсів споробласту й з них формуються полярні капсули; ядра при цьому виштовхуються в порожнину спори, де з часом дегенерують. Дві клітини утворюють зародок з двома гаплоїдними ядрами. У деяких порожнинних видів, що мають невеликі плазмодії з одним-двома вегетативними ядрами та одним-двома генеративними, генеративні безпосередньо перетворюються на споробласти, який розвиваються одна-дві спори, а у великих — тканинних видів — сотні й навіть тисячі спор. Зрілі спори порожнинних міксоспоридій потрапляють у воду через кишечник або нирки, тканинних — через розрив цисти. У деяких видів спори звільняються лише після смерті хазяїна, наприклад, під час паразитування міксоспоридій у хрящовій або нервовій тканині.
У міксоспоридій виробився ряд морфологічних і біологічних адаптацій, що сприяють надходженню паразита в організм хазяїна. Перш за все — це будова спор. У паразитів риб, що харчуються в товщі води, спори мають спеціальні вирости або слизові оболонки, які збільшують їх об'єм і зменшують питому вагу, що допомагає спорам довше зависати у воді. Чим повільніше спускаються спори на дно, тим більше риб ними заражується. Існує також узгодженість між життєвими циклами паразита та хазяїна. У багатьох міксоспоридій період утворення спор припадає на час нересту хазяїна, коли велике скупчення риб на нерестовищах сприяє їх зараженню. Такий тип життєвого циклу трапляється в порожнинних міксоспоридій, що мають невеликі плазмодії й продукують невелику кількість спор.
Види міксоспоридій характерно виділяються розміром і формою спор, що звільняються з хребетних тварин. Наприклад, рід Ceratomyxa є звичайним паразитом жовчного міхура багатьох видів риб, вони мають бумерангоподібні спори з двома полярними капсулами, що нагадує око розміщене в середині спори. Більшість міксоспоридій мають розмір від 10 до 20 мкм, однак Myxidium giganticum сягає до 98 мкм в довжину.
Актиноспоридна стадія
Нещодавно виявлено, що деякі міксоспоридії розвиваються зі зміною хазяїв. Проміжними хазяїнами переважно є кільчасті черви, найчастіше — поліхети. Спори нагадують три або чотири гачки, об'єднані між собою, що є таксономічною ознакою. Так, спори паразита хрящів лососів Myxosoma cerebralis у забрудненій воді активно поглинаються малощетинковими червами — трубковиками з роду Tubifex. Риби заражаються, поїдаючи інвазованих червів. Ці міксоспоридії патогенні для своїх хазяїв: руйнують тканини та органи, в яких паразитують, крім того, часто викликають тяжку загальну інтоксикацію.
Тривалий час вважали, що ці організми належать до окремого класу — Actinosporea, поки на початку 1990-х років експериментально не було підтверджено їх належність до міксоспоридій (за аналізом 16S субодиниці рибосальної РНК).
Значення
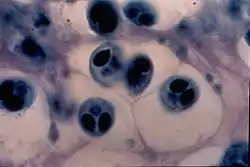
Паразити, які локалізуються в порожнинах внутрішніх органів, як правило, менш патогенні, ніж тканинні. Наприклад, Myxidium ieberkuhni, що паразитує в сечовому міхурі щуки, призводить лише до незначного руйнування стінок міхура. Хоча паразит жовчного міхура форелі Chloromyxus truttae спричинює його різке збільшення, порушення функції й навіть смерть.
Різні види роду Myxobolus заражають зябра та інші органи переважно прісноводних промислових риб. Так, Myxobolus cyprini паразитує в зябрах, м'язах, нирках, печінці, селезінці коропів і викликає злоякісну анемію. Myxobolus pfeifferi паразит м'язів вусача, спричинює переродження м'язів, утворення під шкірою пухлин розмірами з куряче яйце, заповнених безліччю спор і продуктами розпаду тканин. М'ясо риб, уражених цими міксоспоридіями, непридатне для вживання в їжу. Відомі випадки масової загибелі риб від міксоспоридіозу. Myxosoma cerebralis викликає також масову загибель мальків форелі та інших лососевих. Паразит заражає хрящові тканини, в результаті чого порушується їх нормальне функціонування, особливо небезпечним є руйнування капсули навколо органа рівноваги. Хворі риби втрачають вдатність орієнтуватися під час руху й починають крутитися на місці. Спостерігається також викривлення хребта. Великої шкоди завдають міксоспоридії роду Kudoa, що паразитують у багатьох масових видів промислових риб, таких як оселедець атлантичний, мерлуза, путасу. Їх спори мають не дві, а чотири або більше полярних капсули, плазмодій локалізується в м'язовій тканині риб. Під час промислу рибу заморожують і в такому стані зберігають тривалий час, проте навіть за дуже низьких температур міксоспоридії не гинуть і зберігають життєздатність. У разі підвищення температури вони виділяють речовини, що перетворюють м'язи на драглисту масу. Особливо небезпечним патогенний вплив міксоспоридій є в ставкових господарствах, де скупченість риби створює умови для масового зараження й загибелі риб.
Література
Джерела
- Bartholomew, J. L., M. J. Whipple, D. G. Stevens and J. L. Fryer. (1997). The life cycle of Ceratomyxa shasta, a myxosporean parasite of salmonids, requires a freshwater polychaete as an alternate host. American Journal of Parasitology. 83:859-868.
- Wolf, K. & Markiw, M.E. (1984). Biology contravenes taxonomy in the Myxozoa: new discoveries show alternation of invertebrate and vertebrate hosts. Science 225:1449-1452
- Kent, M. L., Margolis. L., & Corliss, J. 0. (1994). The demise of a class of protists: Taxonomic and nomenclatural revisions proposed for the protist phylum Myxozoa Grasse, 1970. Canadian Journal of Zoology 72:932-937.
- Зоологія безхребетних: Підручник: У трьох книгах. Книга 1 / Г. Й. Щербак, Д. Б. Царичкова, Ю. Г. Вервес— К. : Либідь, 1995.—320 с.