Рибосома
Рибосома (ribosome) є немембранною органелою клітини, що складається з рРНК та рибосомних білків (протеїнів). Рибосома здійснює біосинтез білків, транслюючи з мРНК поліпептидний ланцюг. Таким чином, рибосому можна вважати фабрикою, що виготовляє білки, базуючись на наявній генетичній інформації. В клітині дозрілі рибосоми знаходяться переважно в компартментах, для активного білкового синтезу. Вони можуть вільно плавати в цитоплазмі або бути прикріпленими до цитоплазматичного боку мембран ендоплазматичного ретикулуму чи ядра. Активні (ті що є в процесі трансляції) рибосоми знаходяться переважно у вигляді полісом. Існує ряд свідчень, які вказують на те, що рибосома є рибозимом.
Історичний огляд
Рибосоми було виявлено на початку 1950-тих років. Перше глибоке дослідження та опис рибосом, як клітинних органел, було здійснене Джорджем Паладе. За іменем дослідника, рибосоми були названі «частинками Паладе», але згодом, в 1958 році, їх було перейменовано в «рибосоми» з огляду на високий вміст РНК. Роль рибосом в біосинтезі білків було встановлено більш ніж десятиліттям пізніше.
Синтетична рибосома
Після від'єднання від мРНК і початком нового раунду трансляції рибосомальні мала і велика субодиниці відділяються одна від одної. Тому, створення синтетичної рибосоми було технічно складним, оскільки синтетичні та наявні у клітині субодиниці змішувалися від раунду до раунду трансляції[1].
Починаючи з кінця 90х років 20 століття вдалося згенерувати декілька видів мутантних малих субодиниць рибосоми (див. пункт Будова рибосоми), які мали специфічну послідовність у 16S рРНК і з'єднувалися з мРНК, у якої Послідовність Шайна — Дальгарно була специфічно синтезована для взаємодії з модифікованою 16S рРНК. Це дозволило виконувати відбір мутованих малих субодиниць РНК від нативних і інтрудукувати декілька мутацій для вивчення властивостей синтезу білка[1].
Проте велика рибосомальна субодиниця презентувала проблеми, оскільки при створенні синтетичного варіанту не було можливості змусити його не відділятися від мРНК чи від малої субодиниці після завершення одного раунду трансляції[1]. Велика субодиниця містить важливі для вивчення структури, такі як канал для виходу синтезованого білка та сайт PTC (англ. peptidyl transferase centre), в якому відбувається з'єднання амінокислоти, що приєднана до тРНК, яка знаходяться на А-сайті рибосоми, до пептидильного ланцюга, що з'єднаний до молекули тРНК, яка знаходиться на P-сайі рибосоми (див. пункт «Центри зв'язування РНК»)
У липні 2015 року вдалося синтезувати першу повністю синтетичну рибосому. Для того, щоб велика і мала субодиниці не від'єднувались, їх було зв'язано в одну молекулу шляхом синтезу 16S-23S конструкту (Ribo-T). Така синтетична рибосома успішно виконувала синтез білка не лише in vitro, але і підтримувала ріст E.coli при вимушеній відсутності нативних рибосом[1].
Будова рибосоми
Загальна будова
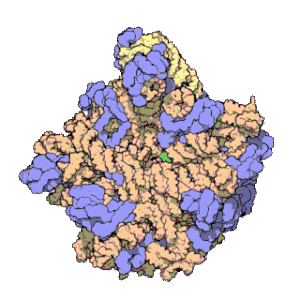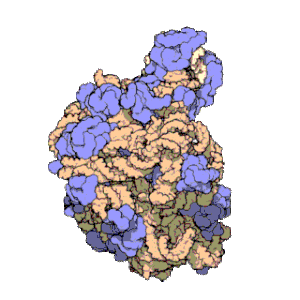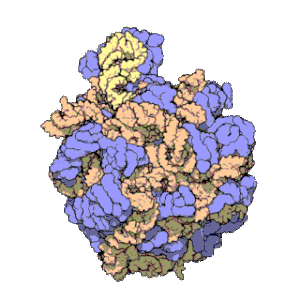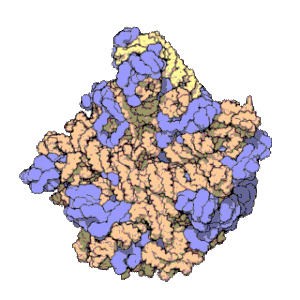
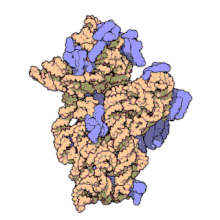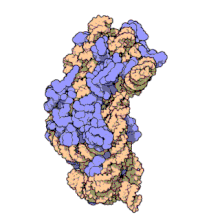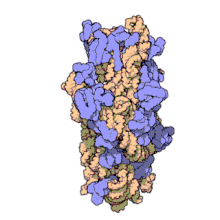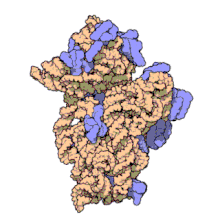
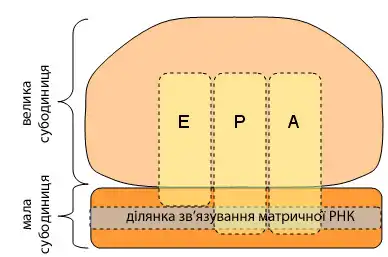
Рибосоми прокаріотів та еукаріотів є дуже подібними за будовою та функцією, але відрізняються розміром. Вони складаються з двох субодиниць: однієї великої та однієї малої. Для процесу трансляції необхідна злагоджена взаємодія обох субодиниць, що разом становлять комплекс із молекулярною масою декілька мільйонів дальтон (Da). Субодиниці рибосом за звичай позначаються одиницями Сведберга (S), що є мірою швидкості седиментації під час центрифугування і залежать від маси, розміру та форми частинки. Позначені в цих одиницях, велика субодиниця є 50S або 60S (прокаріотичні або еукаріотичні, відповідно), мала є 30S або 40S, і ціла рибосома (комплекс малої разом з великою) 70S або 80S.
Молекулярний склад
Молекулярний склад рибосом є доволі складним. Для прикладу, рибосома дріжджів ‘Saccharomyces cerevisiae’ складається із 79 рибосомних білків та 4 різних молекул рРНК. Біогенез рибосом є також надзвичайно складним і багатоступеневим процесом, що відбувається в ядрі та ядерці еукаріотичної клітини.
Атомна структура великої субодиниці (50S) організму Haloarcula marismortui була опублікована N. Ban et al. в журналі Science 11 Серпня 2000 року. Невдовзі після цього, 21шого Вересня 2000 року, B.T. Wimberly, et al., опублікували в журналі Nature структуру 30S субодиниці організму Thermus thermophilus. Використовуючи ці координати, M.M. Yusupov, et al. зуміли реконструювати цілу 70S частинку Thermus thermophilus і опублікувати її в журналі Science, в Травні 2001. У 2009 році професор Джордж Чьорч (George Church) та колеги з Гарварду створили повністю функціональну штучну рибосому в звичайних умовах, які присутні в клітинному середовищі. Як конструкційні елементи використовувались молекули з розщепленої за допомогою ензимів кишкової палички. Створена рибосома успішно синтезує білок, що відповідає за біолюмінесценцію.
Центри зв’язування РНК
Рибосома містить чотири сайти зв'язування для молекул РНК: один для мРНК і три для тРНК. Перший сайт зв’язування тРНК називається сайтом ‘аміноацил-тРНК’, або ‘А-сайтом’. В цьому сайті міститься молекула тРНК „заряджена” „наступною” амінокислотою. Другий сайт, ‘пептидил-тРНК’ зв'язуючий, або ‘P-сайт’, містить молекулу тРНК, що зв’язує ростучий кінець поліпептидного ланцюга. Третій сайт, це ‘сайт виходу’, або ‘E-сайт’. В цей сайт потрапляє порожня тРНК яка позбулась ростучого кінця поліпептида, після його взаємодії з наступною „зарядженою” амінокислотою в пептидильному сайті. Сайт зв’язування мРНК знаходиться в малій субодиниці. Він утримує рибосому „нанизаною” на мРНК котру рибосома транслює.
Функція
Рибосома є органелою, на якій відбувається трансляція генетичної інформації, закодованої в мРНК. Ця інформація втілюється в синтезований тут-же поліпептидний ланцюг. Рибосома несе двояку функцію: є структурною платформою для процесу декодування генетичної інформації з РНК, та володіє каталітичним центром відповідальним за формування пептидного зв'язку, так званим ‘пептидил-трансферазним центром’. Вважається що пептидил-трансферазна активність асоціюється з рРНК, і тому рибосома є рибозимом.
Локалізація рибосом
Рибосоми класифікуються як вільні (перебувають у гіалоплазмі) та невільні або прикріплені (пов'язані з мембранами ендоплазматичної мережі).
Вільні та прикріплені рибосоми відрізняються тільки локалізацією, але вони є структурно ідентичні. Рибосому називають вільною або прикріпленою в залежності від того чи білок який синтезується має ЕР-націлену сигнальну послідовність, тому індивідуальна рибосома може бути прикріпленою створюючи один білок, але вільною в цитозолі коли створює інший білок.
Рибосоми іноді називають органелами, але використання терміну органела обмежується субклітинними компонентами які мають фосфоліпідну мембрану, а рибосома(being entirely particulate) такою не є. Тому рибосоми іноді описують як «немембранні органелли».
Загальна інформація
В еукаріотичних організмах рибосоми можна знайти не лише в цитоплазмі, а й всередині в деяких великих мембранних органелах, зокрема міторибосоми в мітохондріях та специфічні рибосоми хлоропластів. Будова та молекулярний склад цих рибосом відрізняється від складу загально-клітинних рибосом, і є ближчим до складу рибосом прокаріотів. Такі рибосоми синтезують органело-специфічні білки, транслюючи органело-специфічну мРНК.
В еукаріотичних клітинах довгий час вважалося, що рибосоми, прикріплені до ендоплазматичного ретикулуму виконують синтез білків, які будуть секретовані назовні або трансмембранних чи інших сигнальних білків, приєднаних до плазмалеми. Рибонуклеопротеїн SRP (англ. Signal recognition particle) виконує розпізнавання тих білків в процесі синтезу, які повинні бути трансмембранними і приєднує рибосому до ендоплазматичного ретикулуму. Проте останнім часом дослідження вказують, що 50-75 % рибосом можуть бути прикріплені до ЕПР через не до кінця з'ясовані механізми і більшість білків в клітині проходить синтез у рибосомах, які прилягають до ЕПР. Так, у клітинної лінії HEK-293 75 % мРНК відповідяє цитозольним білкам, проте до 50 % рибосом зв'язані з ЕПР[2].
Хвороби
Вважається, що генетичні дефекти рибосомних білків та факторів біогенезу рибосом є летальними на ранніх ембріональних стадіях розвитку вищих організмів. Експериментальний мутагенез рибосомних білків в Drosophila melanogaster (мутації minute) викликає спільний фенотип: занижена швидкість мітозу, зменшений розмір тіла, занижена фертильність, короткі війки. Існує ряд свідчень що пов'язують ракову трансформацію клітин ссавців з розладами трансляційної системи загалом та розладами системи біогенезу рибосом зокрема.
Див. також
Примітки
- Cedric Orelle, Erik D. Carlson, Teresa Szal, Tanja Florin, Michael C. Jewett & Alexander S. Mankin (July 2015). Protein synthesis by ribosomes with tethered subunits. Nature. PMID 26222032. doi:10.1038/nature14862.
- David W. Reid & Christopher V. Nicchitta (April 2015). Diversity and selectivity in mRNA translation on the endoplasmic reticulum. Nature reviews. Molecular cell biology 16 (4): 221–231. PMID 25735911. doi:10.1038/nrm3958.
Використана література
- Alberts, B. 1994. Molecular Biology of the Cell. Garland Pub., New York. xliii, 1294, 67 p.
- Nissen, P., J. Hansen, N. Ban, P.B. Moore, and T.A. Steitz. 2000. The structural basis of ribosome activity in peptide bond synthesis. Science. 289:920-30.
- Ruggero, D., and P.P. Pandolfi. 2003. Does the ribosome translate cancer? Nat Rev Cancer. 3:179-92.
- Sydorskyy, Y. 2005. Identification and characterization of novel ribosome biogenesis factors. Doctoral Thesis, Department of Cell Biology, University of Alberta. Edmonton, Canada.
- Uechi, T., T. Tanaka, and N. Kenmochi. 2001. A complete map of the human ribosomal protein genes: assignment of 80 genes to the cytogenetic map and implications for human disorders. Genomics. 72:223-30.
- Venema, J., and D. Tollervey. 1999. Ribosome synthesis in Saccharomyces cerevisiae. Annu Rev Genet. 33:261-311.