Нуклеація елементів цитоскелету
Нуклеація — це початковий і лімітуючий етап полімеризації елементів цитоскелету, що полягає у формуванні стабільного ядра із кількох мономерів. Вільні субодиниці актину чи тубуліну у розчині можуть взаємодіяти між собою і утворювати нековаленті зв'язки, але такі комплекси зазвичай швидко дисоціюють, і для їх стабілізації необхідне приєднання більшої кількості мономерів. Якщо це відбудеться, то стабільне угрупування виступатиме ядром для полімеризації наступних молекул. У пробірці процес утворення ядра може тривати досить довго, але в живій клітині є спеціальні системи, що каталізують нуклеацію мікротрубочок та мікрофіламетів.
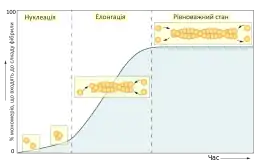
Нуклеація мікротрубочок у клітині відбувається у спеціальних структурах, що мають назву центри організації мікротубочок (ЦОМТ), і потребує участі γ-тубуліну. Нуклеація мікрофіламентів зазвичай має місце поблизу клітинної мембрани, цей процес відбувається за участі одної з трьох систем: комплексу ARP 2/3, білків формінів або білків Spire. Про динаміку полімеризації проміжних філаментів у клітині відомо менше.[1]
Нуклеація мікротрубочок
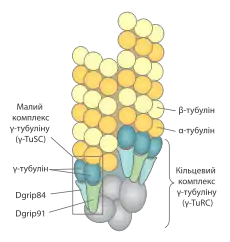
Мікротурбочки починають полімеризвуватись у ЦОМТ, де відбувається їх нуклеація на (-)кінці. Для цього процесу необхідний γ-тубулін, консервативний білок на 30 % ідентичний за амінокислотною послідовністю до α- та β-тубуліну, та інші білки, що формують разом із ним кільцевий комплекс γ-тубуліну (англ. γ-Tubulin Ring Complex, γ-TuRC). Комплекси γ-TuRC у тваринній клітині є частиною перицентріолярної речовини центросом, тоді як у рослин та грибів він асоційований із ядерною оболонкою.[1]
Основними функціями γ-TuRC крім каталізу самого процесу нуклеації, є те, що вони стабілізують (-)кінці та слугують зразком для збирання мікротрубочок. Мікротрубочки, що полімеризуються in vitro можуть мати різну кількість протофіламентів (в середньому 14), тоді як в утворених in vivo їх кількість завжди рівна 13, це важливо, тому що тільки в такому випадку протофіламенти розміщуються строго паралельно один до одного і не утворюють спіралей, а це полегшує везикулярний транспорт вздовж них.[2]
Складовою частиною γ-TuRC є малі комплекси γ-тубуліну (англ. γ-Tubulin Small Complex, γ-TuSC), що побудовані із двох молекул самого γ-тубуліну та ще двох білків: Dgrip84 та Dgrip91 (англ. Drosophila gamma ring protein) у дрозофіли, Spc97 та Spc98 у дріжджів, Xgrip109 та Xgrip110 у Xenopus laevis[3] та GCP2 і GCP3 у людини.[4] Вони мають V-подібну форму та розміщуються по спіралі у складі γ-TuRC. Кожна молекула γ-тубуліну в такому колі взаємодіє із α-тубуліном у димерній субодиниці мікротрубочок. Проте, оскільки γ-TuSC мітить 2 молекули γ-тубуліну, їх кількість мусить бути парною, а отже не може бути рівною кількості протофіламентів у мікротрубочці. Швидше за все, γ-TuRC містить шість комплексів γ-TuSC (а отже 12 γ-тублінів), в одному місці між ними існує проміжок. Крім γ-TuSC до складу кільцевого комплексу також входять інші допоміжні білки, що утворюють кеп («шапочку») на його кінці. Кеп може брати участь у прикріпленні всієї структури до центросоми або виконувати інші функції.[3]
Нуклеація мікрофіламентів
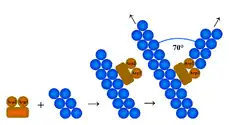
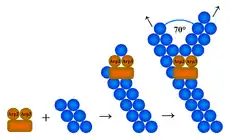
Так як більшість із мікрофіламентів у клітині розміщуються у клітинному кортексі (тонкому шарі цитоплазми відразу ж під плазматичою мембраною), там і відбувається їхня нуклеація. Для нуклеації актинових філаментів необхідне утворення ядра із трьох мономерів, каталізувати цей процес можуть три різні за механізмом дії системи: комплекс ARP 2/3 (англ. Actin Related Protien), білки форміни та білок Spire.[1][5]
Комплекс ARP 2/3
Комплекс ARP 2/3 каталізує нуклеацію на (-)кінці актинових філаментів, при чому він забезпечує також і їх галуження під кутом 70°. Він складається із семи субодиниць, за двома із яких: ARP2 та ARP3, ця структура і отримала свою назву. Білки ARP2 та ARP3 приблизно на 45 % ідентичні актину. Решта п'ять субодиниць називаються ARPC1-5. Механізм дії комплексу ARP 2/3 полягає у тому, що білки ARP2 та ARP3 імітують димер актину, і після приєднання ще однієї молекули актину стають ядром полімеризації. Проте сам по собі ARP 2/3 проявляє мало активності, імовірно, через те, що білки ARP2 та ARP3 не перебувають у потрібній конформації. Зміна їхньої просторової укладки може відбутись під впливом факторів, що стимулюють нуклеацію (англ. Nucleation promoting factors, NPF) пілся, чого комплекс ефективно каталізує утворення нового філаменту. У клітинах існує багато різних білків NPF, всі вони регулюються сигнальними шляхами, що впливають на актиновий цитоскелет.
Існують дві моделі, що описують механізм галуження мікрофіламентів за участі комплексу ARP2/3:
- Галуження збоку (англ. side end branching): комплекс ARP 2/3 приєднується до боку материнського філаменту і каталізує утворення дочірнього під кутом 70°; цю гіпотезу підтверджує більша кількість експериментальних даних;
- Галуження з (+)кінця (англ. barbed end branching): комплекс ARP 2/3 приєднується до плюс кінця материнського філаменту і каталізує утворення двох гілок, розміщених під кутом 70° одна до одної.
Комплекс ARP2/3 важливий для утворення багатьох клітинних структур, таких як ламелоподії, філоподії, а також для процесу фагоцитозу.[5]
Форміни
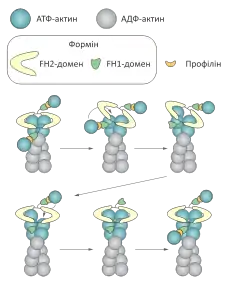
Деякі структури в клітині потребують не розгалуженої сітки, утворення якої забезпечує комплекс Arp 2/3, а із пучків паралельно розташованих мікрофіламентів. В такому випадку нуклеація часто відбувається за участі білків класу формінів. На відміну від комплексу Arp 2/3 вони асоціюються із (+)кінцем актинового філаменту і пришвидшують його ріст. Форміни залишаються зв'язаними із (+)кінцем під час процесу полімеризації, і не дозволяють приєднуватись іншим регуляторним білкам, які можуть її термінувати.[1]
Форміни присутні майже у всіх досліджених еукаріотичних клітинах. Їхньою характерною ознакою є наявність двох доменів гомології формінів (англ. Formin Homology domain) — FH1 та FH2. Безпосередню участь у нуклеації бере домен FH2, і його достатьно для цього процесу. Форміни активні як гомодимери, у складі яких їхні FH2 домени утворюють кільце, що охоплює дві молекули актину. Така конформація білка називається «закритою» і вона перешкоджає додаванню нових субодиниць до мікрофіламенту. Проте формін може переходити у «відкриту» конформацію, яка сприяє полімеризації на (+)кінці: існують дві гіпотези, що описують цей процес. Згідно із першою формін спочатку просувається на один «крок» вздовж мікрофіламенту звільняючи місце для приєднання нової молекули актину, згідно із другою, навпаки — спочатку приєднується новий актиновий мономер, після чого формін посувається на один «крок».[6]
FH1 домен не є необхідним для процесу нуклеації за участі формінів, але пришвидшує цей процес, як і полімеризацію. Він містить багаті на пролін послідовності, які забезпечують зв'язування із профіліном — білком доставки мономерного актину. Домени FH1 знаходяться на довгих гнучких «вусах» молекули форміну, і коли вона переходить у відкриту конформацію, ці вуса можуть згинатись і доставляти мономер актину, приєднаний до профіліну точно на його місце у мікрофіламенті. Після цього профілін дисоціює, а домен FH1 може «навантажуватись» наступними комплексами профілін-актин.[1][6]
Білок Spire
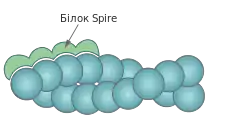
Нещодавно було виявлено ще один клас білків, що забезпечують нуклеацію мікрофіламентів — родину WH2 нуклеаторів, характерною ознакою яких є наявність кількох доменів WH2 (англ. WASP Homology 2), які також наявні і в деяких NPF (білків, що активують комплекс ARP 2/3). Одним із найкраще вивчених білків цієї родини є Spire ссавців.
Білок Spire містить чотири домени WH2, кожен з яких може взаємодіяти із мономерним актином. Таким чином він може забезпечувати нуклеацію, утворюючи і стабілізуючи ядро із чотирьох мономерів. Проте деталі цього механізму залишаються нез'ясованими: деякі дослідження вказують на те, що Spire асоціює із (-)кінцем, тоді як інші свідчать про взаємодію із (+)кінцем; також незрозуміле взаємне розміщення актинових мономерів у тетрамерному ядрі під час нуклеації.[6]
Джерела
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Kollman JM, Polka JK, Zelter A, Davis TN, Agard DA. (2010). Microtubule nucleating gamma-TuSC assembles structures with 13-fold microtubule-like symmetry. Nature 466: 879–82. PMID 20631709. doi:10.1038/nature09207.
- Moritz M, Agard DA. (2001). Gamma-tubulin complexes and microtubule nucleation. Curr Opin Struct Biol 11: 174–81. PMID 11297925. doi:10.1016/S0959-440X(00)00187-1.
- Moritz M, Braunfeld MB, Guénebaut V, Heuser J, Agard DA (2000). Structure of the gamma-tubulin ring complex: a template for microtubule nucleation. Nat Cell Biol 2: 365–70. PMID 10854328. doi:10.1038/35014058.
- Goley ED, Welch MD (2006). The ARP2/3 complex: an actin nucleator comes of age. Nat Rev Mol Cell Biol 7: 713–26. PMID 16990851. doi:10.1038/nrm2026.
- Campellone KG, Welch MD (2010). A nucleator arms race: cellular control of actin assembly. Nat Rev Mol Cell Biol. 11 (4): 237–51. PMID 20237478. doi:10.1038/nrm2867.
Посилання
- MBInfo: Microtubule (англ.)
- MBInfo: Tubulin Complex Assembly (англ.)