Мікротрубочки
Мікротрубочки — білкові внутрішньоклітинні структури, що входять до складу цитоскелету еукаріотів.
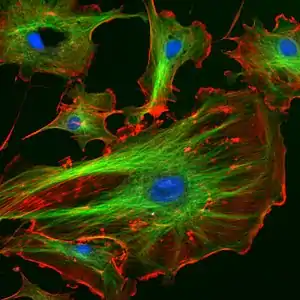
Мікротрубочки є циліндрами діаметром 25 нм з порожниною усередині. Їх довжина може бути від кількох мікрометрів до, ймовірно, кількох міліметрів (в аксонах нервових клітин). Їх стінка утворена димерами тубуліну. Мікротрубочки, подібно актиновим мікрофіламентам, полярні: на одному кінці відбувається самозбирання мікротрубочки, на іншому — розбирання. У клітинах мікротрубочки грають роль структурних компонентів і беруть участь в багатьох клітинних процесах, включаючи мітоз, цитокінез і везикулярний транспорт.
Будова
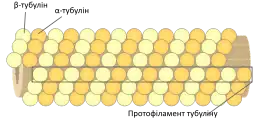

Мікротрубочки — структури, в яких 13 тубулінових α- та β-гетеродимерів укладені по колу полого циліндра. Зовнішній діаметр циліндра становить близько 25 нм, внутрішній, — близько 15 нм. Один з кінців мікротрубочки, що називається позитивним кінцем (або плюс-кінцем), постійно приєднує до себе вільний тубулін. Від протилежного кінця — негативного (мінус-кінця) — тубулінові субодиниці відщеплюються.
В утворенні мікротрубочки in vitro виділяють три фази:
- сповільнена фаза, або нуклеація. Це етап зародження мікротрубочки, коли молекули тубуліна починають з'єднуватися в крупніші утворення. Таке з'єднання відбувається повільніше, ніж приєднання тубуліна до вже зібраної мікротрубочки, тому фаза і називається сповільненою.
- фаза полімеризації, або елонгація. Якщо концентрація вільного тубуліну висока, його полімеризація відбувається швидше, ніж деполімерізация на негативному кінці, за рахунок чого мікротрубочка подовжується. У міру її зростання концентрація тубуліну падає до критичної і швидкість росту сповільнюється до вступу до наступної фази;
- фаза стабільного стану. Деполімерізация врівноважує полімеризацію, і зростання мікротрубочки зупиняється. Лабораторні дослідження показують, що збірка мікротрубочок з тубуліну відбувається тільки в присутності гуанозинтрифосфату та іонів магнію, оптимально при температурі 37 °C.
Динамічна нестабільність
Мікротрубочки є динамічними структурами і в клітині постійно полімеризуються і деполімерізуются. Центросома, локалізована поблизу ядра, виступає як центр організації мікротрубочок: вони ростуть від неї до периферії клітини. В той же час мікротрубочки можуть раптово припинити своє зростання, у цьому разі вони зазвичай швидко скорочуються назад у напрямку до центросоми до повного розчинення, а потім виростають знову. Мінус-кінець мікротрубочки зазвичай закріплений в центросомі.
Динамічна нестабільність мікротрубочок грає важливу фізіологічну роль. Наприклад, при поділі клітини мікротрубочки ростуть дуже швидко і сприяють правильній орієнтації хромосом і утворенню веретена поділу.
Функція
Мікротрубочки в клітині використовуються як канали для транспортування «вантажів». Транспортування по мікротрубочках здійснюють білки, що називаються моторними. Це високомолекулярні сполуки, що складаються з двох важких (масою близько 300 кДа) і декількох легких ланцюгів. У важких ланцюгах виділяють головний і хвостовий домени. Два головні домени зв'язуються з мікротрубочками і є власне двигунами, а хвостові — зв'язуються з органелами та іншими внутріклітинними утвореннями, що підлягають транспортуванню.
Виділяють два види моторних білків, здатних рухатися уздовж мікротрубочок:
- цитоплазматичні динеїни переміщують «вантаж» тільки від плюс-кінця до мінус-кінця мікротрубочки, тобто з периферійних областей клітини до центросоми;
- кінезини, переміщуються до плюс-кінця, тобто до клітинної периферії.
Переміщення здійснюється за рахунок енергії АТФ. Головні домени моторних білків для цього містять АТФ-зв'язуючі ділянки. Крім транспортної функції, мікротрубочки формують центральну структуру війок і джгутиків еукаріотів — аксонему. Типова аксонема містить 9 пар об'єднаних мікротрубочок і дві повні мікротрубочки.
Рослинні мікротрубочки
Мікротрубочки рослин є високодинамічними складовими цитоскелету, які залучені до важливих клітинних процесів, зокрема, сегрегації хромосом, формування фрагмопласту, мікрокомпартменталізації, внутрішньоклітинного транспорту, а також до підтримання сталої форми та полярності клітини. Мобільність мікротрубочок забезпечується динамічною нестабільністю, пересуванням полімерів моторними білками, тредмілінгом та гібридним механізмом тредмілінгу із динамічною нестабільністю плюс-кінця та повільною деполімеризацією мінус-кінця [1].
Організація та динаміка
(внизу, у центрі).
Мікротрубочки є надмірно чутливими до біотичних та абіотичних факторів навколишнього середовища (холоду, освітлення, посухи, засолення, впливу гербіцидів і пестицидів, затоплення, стискання, впливу електричного поля, тиску та сили тяжіння), а також фітогормонів, антимітотичних препаратів та ряду інших біологічно активних сполук [2]. Мікротрубочки є порожнистими полярними циліндричними філаментами діаметром понад 24 нм, що збираються з гетеродимерів α- та β-тубулінів, які у положенні «голова-до-хвоста» формують 13 протофіламентів.
Суттєве обмеження імуногістохімічних досліджень полягає у неможливості прижиттєвої візуалізації динаміки мікротрубочок еукаріотичних та прокаріотичних клітин в режимі реального часу. Це обмеження було подолано завдяки застосуванню конфокальної мікроскопії із зеленим флуоресцентним білком GFP, ізольованим із медузи Aequorea victoria L. [3]. Репортерна конструкція GFP-MBD для гетерологічної трансформації навіть за низького рівня транзієнтної експресії in vivo та in vitro дозволяє візуалізувати динамічну нестабільність мікротрубочок в різних типах клітин кореня. [4] [5]
У клітинах вищих рослин присутні чотири типи побудов мікротрубочок:
- сітка кортикальних та ендоплазматичних мікротрубочок,
- препрофазна стрічка,
- веретено поділу,
- фрагмопласт [6].
Білки, асоційовані з мікротрубочками
Усі компоненти цитоскелету та інші органели пов'язані між собою низкою специфічних білків, асоційованих з мікротрубочкми (БАМ). У тваринних клітинах найбільш дослідженими БАМ є tau і БАМ2, які стабілізують мікротрубочки та приєднують їх до інших клітинних структур, а також транспортні білки динеїн та кінезин [7]. Функціонування різних груп рослинних мікротрубочок залежить від наявності ізоформ БАМ з родини БАМ65 та регуляторних кіназ і фосфатаз. Зокрема, висококонсервативний тваринний гомолог родини БАМ65 є суттєвим для набуття мікротрубочками певних конфігурацій протягом розвитку рослини. [5] Орієнтація й організація різних популяцій та типів побудов мікротрубочок є ткане- та органоспецифічною [8].
Побудова кореня різушки Таля Arabidopsis thaliana L. є типовою для дводольних рослин. Найближчим до поверхні кореня є епідермальний шар, клітини якого у зрілій зоні в залежності від здатності ініціювати розвиток кореневих волосків є трихобластами чи атрихобластами [9]. Глибше розташовані накопичувальний безхлоропластний кортикальний шар із численними міжклітинниками та плазмодесмами та шар ендодермальних клітин із поясками Каспарі на антиклінальних поверхнях [10]. Центральний циліндр кореня формують паренхімні клітини перициклу [10], які здатні до швидких поділів, та елементи ксилеми і флоеми. Присутнє й функціональне розмежування кореневих зон: зони поділу, елонгації, дозрівання, а також перехідна зона на межі зон ініціації та елонгації. З перициклу формуються бічні корені, а з трихобластів епідермального шару — кореневі волоски [11] [10]. Кінчик кореня вкритий кореневим чохликом із специфічною морфологією клітин колумели.
Кортикальні мікротрубочки
Ацентросомальні кортикальні мікротрубочки (КМТ) важливі для морфогенезу рослин, регуляції клітинного поділу і елонгації [12]. Високодинамічна популяція мембранозв'язаних коротких КМТ швидко реорієнтується з інтерфазного поперечного положення у поздовжнє при елонгації клітини [13]. Ацентросомальні кортикальні мікротрубочки мають невпорядковане розміщення плюс-кінців і виявляють динамічну нестабільність, а вільні мінус–кінці КМТ повільно деполімеризуються, тобто КМТ самоорганізуються гібридним механізмом динамічної нестабільності та тредмілінгу [1]. Енуклеація відбувається по всій поверхні плазматичної мембрани [13] [1]. Білок SPR1 регулює динаміку та організацію плюс-кінця КМТ рослин, що позначається на анізотропному рості клітини [14] [15]. Ацентросомальні кортикальні мікротрубочки розташовуються паралельно до целюлозних мікрофібрил [16], правильна організація КМТ є суттєвою для нормального синтезу клітинної стінки [17]. Встановлено, що КМТ об'єднуються у вузли, які часто перетинаються для стабілізації мікротрубочок та втримання білків на їх поверхні [15].
Латеральні циліндричні вирости трихобластів, кореневі волоски, досягають значної довжини відносно власної товщини з досить сталим діаметром у Arabidopsis thaliana L.: (незрілі ~ 6–10 нм; зрілі — понад 1 мм) і характеризуються високополярною цитоархітектурою [18]. Видовження їх відбувається за допомогою верхівкового росту (англ. tip growth) шляхом поляризованого екзоцитозу, що відзначається зворотно-фонтанним током цитоплазми, градієнтом цитоплазматичного Ca2+, активністю F-актину та зміщенням клітинного вмісту до верхівки волоска. На ранніх стадіях розвитку кореневі волоски 3-денних проростків Arabidopsis thaliana L. ростуть зі швидкістю 0,4 мкм/хв, прискорюючись пізніше до 1—2,5 мкм/хв [18].
Рослинним клітинам притаманна організована популяція кортикальних мікротрубочок [7], яка у кореневих волосках присутня на усіх рівнях розвитку [19] [20]. При переході із зародкового стану до стану видовження, кортикальні мікротрубочки верхівки волосків не візуалізуються, оскільки з'являються ендоплазматичні мікротрубочки. Кортикальні мікротрубочки орієнтовані поздовжньо або спірально [20] [21]. У кукурудзи Zea mays L. та салату Lactuca sativa L. ініціація росту кореневих волосків пов’язана з реорганізацією популяції КМТ у трихобластах [22] [23] [20]. Ця популяція контролює стабільність та напрямок апікального росту кореневих волосків [24] [25]. Порівняння чотирьох стандартних параметрів динамічної нестабільності КМТ in vivo — рівня ростової активності, швидкості розбирання, частоти переходів від розбирання до росту («порятунок») і навпаки («катастрофа») виявило, що КМТ молодих кореневих волосків є динамічнішими за зрілі. Сітка мікротрубочок реорганізується у відповідь на змінні параметри навколишнього середовища та стимули диференціації шляхом варіювання показників динамічної нестабільності [25].
Примітки
- Shaw et al., 2003.
- Weber and Westermann, 2003.
- Ueda, 1999.
- Marc et al., 1998.
- Wasteneys and Yang, 2004.
- Barlow and Balushka, 2000.
- Goddard et al., 1994.
- Lloyd, 1994.
- Sugimoto et al., 2000.
- Dolan et al., 1993.
- Рейвн и др., 1990.
- Dixit et al., 2006.
- Yuan et al., 1994.
- Dixit and Cyr , 2004.
- Lloyd., 1994.
- Baskin et al., 2004.
- Burk et al., 2006.
- Dolan et al., 1994.
- Sieberer et al., 2002.
- Van Bruaene et al., 2004.
- Sieberer et al., 2005
- Balusˇka et al., 2000.
- Geitmann and Emons, 2000.
- Bibikova et al., 1999.
- Vassileva et al., 2005.
Література
- Фаллер Дж. М., Шилдс Д., Молекулярная биология клетки. Руководство для врачей. Пер. с англ. — М.: «БИНОМ», 2006. — 256 с. (рос.)
