Дріжджі
Дрі́жджі — позатаксономічна група одноклітинних грибів, які втратили міцеліальну будову внаслідок переходу до проживання у рідких і напіврідких субстратах, багатих на органічні речовини. Об'єднує більше 2000 [1] [2] видів , що належать до аскоміцетів і базидіоміцетів[3] та домінують серед грибів у водних середовищах[4].
? Дріжджі | ||||||
|---|---|---|---|---|---|---|
 | ||||||
| Біологічна класифікація | ||||||
| ||||||
Типовий поділ (парафілетична група) | ||||||
| ||||||
Посилання | ||||||
| ||||||
Межі групи окреслені нечітко: багато грибів, здатних розмножуватися вегетативно в одноклітинній формі, що ідентифікуює їх як дріжджі, на інших стадіях життєвого циклу утворюють розвинений міцелій, а в низці випадків і макроскопічні плодові тіла. Раніше такі гриби виділяли в окрему групу дріжджеподібних, але зараз їх усіх зазвичай розглядають разом із дріжджами[5]. Дослідження 18S рРНК показали близьку спорідненість із типовими грибами інших видів, здатними до росту тільки у вигляді міцелію.
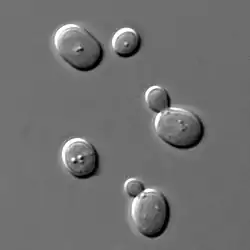
Розміри дріжджових клітин зазвичай становлять 3—7 мкм у діаметрі, водночас деякі види здатні зростати до 40 мкм[6]. Більшість видів розмножується безстатево за допомогою брунькування, хоча деякі види здатні до симетричного (бінарного) поділу.
Дріжджі мають велике практичне значення, особливо пекарські чи пивні дріжджі (Saccharomyces cerevisiae). Деякі види є факультативними патогенами.
Крім того, деякі види надзвичайно важливі як модельні організми у клітинній біології. Дріжджі Saccharomyces cerevisiae та Schizosaccharomyces pombe є найбільш дослідженими еукаріотичними мікроорганізмами. Такі дослідження проводять з метою збору інформації про біологію еукаріотичних клітин і, урешті-решт, про біологію людини[7]. Інші види дріжджів, наприклад Candida albicans, є опортуністичними патогенами і можуть спричиняти інфекції у людини. Зараз повністю розшифрований геном дріжджів Saccharomyces cerevisiae і Schizosaccharomyces pombe (клас схізосахароміцети) — вони стали першими еукаріотами, чий геном був повністю секвенований.
Історія
Слово «дріжджі» має спільний корінь зі словом «дрижати», яким описується спінювання рідини, що часто супроводжує бродіння, здійснюване дріжджами.
Дріжджі, ймовірно, одні з найстародавніших «домашніх організмів». Тисячі років люди використовували їх для ферментації і випічки. Археологи знайшли серед руїн давньоєгипетських міст жорна і пекарні, а також зображення пекарів і пивоварів. Припускають, що пиво єгиптяни почали варити за 6000 років до н. е., а до 1200 року до н. е. опанували технологію випічки дріжджового хліба разом з випічкою прісного[8]. Для початку зброджування нового субстрату люди використовували залишки старого. У результаті, у різних господарствах сторіччями відбувалася селекція дріжджів і сформувалися нові штами, що не трапляються у природі, багато з яких навіть помилково були описані як окремі види. Вони є такими ж продуктами людської діяльності, як сорти культурних рослин.

У 1680 році голландський натураліст Антоні ван Левенгук вперше побачив дріжджі в оптичний мікроскоп, проте через відсутність руху не розпізнав у них живі організми[9]. І лише 1857 року французький мікробіолог Луї Пастер у праці «Mémoire sur la fermentation alcoholique» довів, що спиртове бродіння — не просто хімічна реакція, як вважалося раніше, а біологічний процес, що проводиться дріжджами[10][11].
У 1881 році Еміль Хрістіан Хансен виділив чисту культуру дріжджів, а 1883 року вперше використав її для отримання пива замість нестабільних заквасок[8]. Наприкінці 19 ст. за його участі створюється перша класифікація дріжджів, на початку 20 ст. з'являються визначники і колекції дріжджових культур. У другій половині століття наука про дріжджі (зимологія), окрім практичних завдань, починає приділяти увагу екології дріжджів у природі, клітинній біології, генетиці.
До середини 20 ст. вчені спостерігали тільки статевий цикл аскоміцетних дріжджів і розглядали їх усіх як відособлену таксономічну групу сумчастих грибів (аскоміцетів). Японському мікологу Ісао Банно 1969 року вдалося індукувати статевий цикл розмноження Rhodotorula glutinis, який є базидіоміцетом. Сучасні молекулярні дослідження засвідчили, що дріжджі (одноклітинні форми) сформувалися незалежно серед аскоміцетних і базидіоміцетних грибів і є не єдиним таксоном, а морфологічною формою.
24 квітня 1996 року було оголошено, що Saccharomyces cerevisiae став першим еукаріотичним організмом, чий геном (12 млн пар основ) був повністю секвенований[12]. Секвенування, в якому брали участь понад 100 лабораторій, зайняло 7 років[13]. Наступним дріжджовим організмом і шостим еукаріотом з повністю розшифрованим геномом 2002 року став Schizosaccharomyces pombe[14] з 13,8 млн пар основ.
Біологія дріжджів
Класифікація
Вирізнити дріжджі, що належать до різних відділів грибів, можна як за характеристиками їхнього життєвого циклу, так і без його спостереження — за фізіологічними ознаками. До них належать: синтез каротиноїдів (трапляється лише у базидіоміцетних дріжджів), тип убіхінонів (з 5—7 ізопреноїднимі залишками в аскоміцетних і з 8—10 у базидіоміцетних, хоча є винятки), тип поділу (брунькування або симетричний поділ, див. розділ Життєвий цикл), вміст ГЦ в ДНК (26—48% у аскоміцетних, 44—70% у базидіоміцетних), наявність уреази (характерна за декількома винятками тільки базидіоміцетним) та ін.
Особливості метаболізму
Дріжджі є хемоорганогетеротрофами та використовують органічні сполуки і для отримання енергії, і як джерело вуглецю. Їм необхідний кисень для дихання, проте за його видсутності багато видів здатні отримувати енергію за рахунок анаеробного дихання (бродіння) з виділенням спиртів, тобто вони є факультативними анаеробами. На відміну від бактерій, серед дріжджів немає облігатних анаеробів, що гинуть за наявності кисню в середовищі. Якщо пропускати повітря через зброджуваний субстрат, то дріжджі припиняють бродіння і починають дихати (оскільки цей процес ефективніший), споживаючи кисень і виділяючи вуглекислий газ. Це прискорює зростання дріжджових кліток (ефект Пастера). Проте навіть за доступу кисню у разі високого вмісту глюкози в середовищі дріжджі починають її зброджувати (ефект Кребтрі).
Дріжджі відносно вимогливі до умов середовища. В анаеробних умовах дріжджі можуть використовувати як джерело енергії тільки вуглеводи, причому в основному гексози і побудовані з них олігосахариди. Деякі види (Pichia stipitis, Pachysolen tannophilus) засвоюють і пентози, наприклад ксилозу. Schwanniomyces occidentalis і Saccharomycopsis fibuliger здатні зброджувати крохмаль, Kluyveromyces fragilis — інулін. В аеробних умовах коло засвоєних субстратів ширше: крім вуглеводів, також жири, вуглеводні, ароматичні й одновуглецеві сполуки, спирти, органічні кислоти. Значно більше видів здатні використовувати пентози в аеробних умовах. Проте складні сполуки (лігнін, целюлоза) для дріжджів недоступні.
Джерелами азоту для всіх дріжджів можуть бути солі амонію, приблизно половина видів має нітратредуктазу і може засвоювати нітрати. Шляхи засвоєння сечовини різні в аскоміцетних і базидіоміцетних дріжджів. Аскоміцетні спочатку карбоксилюють її, потім гідролізують, базидіоміцетні — відразу гідролізують уреазою.
Для практичного застосування важливі продукти вторинного метаболізму дріжджів, що виділяються в малих кількостях у середовище: сивушні масла, ацетоїн, діацетил, масляний альдегід, ізоаміловий спирт, диметилсульфід та ін. Саме від них залежать органолептичні властивості отриманих за допомогою дріжджів продуктів.
Життєвий цикл
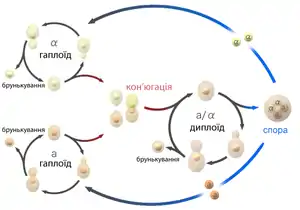
Характерною особливістю дріжджів є здатність до вегетативного розмноження в одноклітинному стані (анаморфи). У порівнянні з життєвими циклами інших грибів це виглядає як брунькування спор або зиготи, або як симетричний (бінарний) поділ. Багато дріжджів здатні також до реалізації примітивного статевого життєвого циклу, в якому можуть бути і міцеліальні стадії (телеоморфи).
У деяких дріжджеподібних грибів, що утворюють міцелій, можливий його розпад на клітини (артроспори). Це відбувається в родах Endomyces, Galactomyces, Arxula і Trichosporon. В останніх двох артроспори після утворення починають розмножуватися брунькуванням. Trichosporon також утворює вегетативні ендоспори всередині клітин міцелію.
Цикл аскоміцетних дріжджів
Найхарактернішим типом вегетативного розмноження для одноклітинних аскоміцетних дріжджів є брунькування, лише Schizosaccharomyces pombe та пов'язані види розмножуються не брунькуванням, а бінарним поділом[15]. Місце утворення бруньки є важливою діагностичною ознакою: полярне брунькування за рахунок утворення шрамів брунькування зумовлює формування апікулярних лимоноподібних (Saccharomycodes, Hanseniaspora, Nadsonia) і грушеподібних (Schizoblastosporion) клітин, тоді як багатобічне брунькування не змінює форму клітини (Saccharomyces, Pichia, Debaryomyces, Candida). У родів Sterigmatomyces, Kurtzmanomyces і Fellomyces брунькування відбувається на довгих виростах (стерігмах).
Брунькування у аскоміцетних дріжджів голобластичне: клітинна стінка материнської клітини розм'якшується, вигинається назовні та дає початок клітинній стінці дочірньої клітини. Часто, особливо у аскоміцетних дріжджів родів Candida і Pichia, клітини після брунькування не розходяться і утворюють псевдоміцелій, що відрізняється від справжнього міцелію виразно видимими перетяжками на місці септ і коротшими порівняно з попередніми кінцевими клітинами.
Гаплоїдні аскоміцетні дріжджові клітини мають два типи спаровування: а і α. Термін «стать» не використовується, оскільки клітки морфологічно ідентичні та відрізняються лише одним генетичним локусом MAT (від англ. mating — спаровування). Клітини різних типів можуть зливатися і утворювати диплоїд a/α, який після мейозу дає 4 гаплоїдних аскоспори: дві а і дві α. Вегетативне розмноження аскоміцетних дріжджів можливе у різних видів або тільки на гаплоїдній стадії, або тільки на диплоїдній, або на обох (гапло-диплоїдні дріжджі).
Цикл базидіоміцетних дріжджів
Брунькування базідіоміцетних дріжджів ентеробластичне: клітинна стінка материнської клітини розривається, з розриву виходить брунька і синтезує свою клітинну стінку з нуля. Бінарний поділ дріжджових клітин для базидіоміцетів не характерний.
Крім звичайного брунькування, багато видів винятково базидіоміцетних дріжджів (роди Sporidiobolus, Sporobolomyces, Bullera) здатні утворювати вегетативні балістоспори: спори на вирості, наповненому глікогеном. Унаслідок гідролізу глікогену тиск збільшується і спора відстрілюється на відстань до декількох міліметрів. Під час тесту на утворення балістоспор дріжджі висіваються на пластинку агарізованного поживного середовища, закріплену на кришці чашки Петрі. Ріст дріжджів на середовищі під цією пластинкою означає наявність у них балістоспор та їхню належність до базидіоміцетів.

У випадку статевого розмноження у базидіоміцетів при злитті гаплоїдних дріжджових клітин (плазмогамія) злиття ядер (каріогамія) не відбувається і формується дікаріотична клітина, що дає початок міцелію. Вже у міцелії відбувається каріогамія і утворюються базидіоспори, часто навіть на плодовому тілі (ряд Tremallales). Єдиними дріжджами серед базидіоміцетів, що не формують міцелій навіть при статевому циклі розмноження, є Xanthophyllomyces dendrorhus.
Слід зазначити, що у базидіоміцетових дріжджів типи спаровування відрізняються зазвичай не одним, а великою кількістю локусів. Можуть зливатися тільки ті клітини, в яких усі ці локуси різні, тобто типів спаровування більше двох.
Поширення у природі
Місця проживання дріжджів пов'язані переважно з багатими цукрами субстратами: поверхнею плодів і листя, де вони харчуються прижиттєвими виділеннями рослин, нектаром квітів, раневими соками рослин, мертвою фітомасою тощо, проте вони поширені також у ґрунті (особливо в підстилці і органогенних горизонтах) і природних водах. Дріжджі (роди Candida, Pichia, Ambrosiozyma) постійно наявні в кишечнику і ходах ксилофагів (комах, що харчуються деревиною), багаті дріжджові співтовариства розвиваються на листі, ураженому попелицею. Представники роду Lypomyces є типовими мешканцями ґрунту.
Застосування
Деякі види дріжджів здавна використовуються людиною у приготуванні алкогольних напоїв (пива, вина, квасу та ін.) і хліба. У поєднанні з перегонкою, процеси бродіння лежать в основі виробництва і міцних спиртних напоїв. Корисні фізіологічні властивості дріжджів дозволяють використовувати їхні біотехнології. У наш час[коли?] їх застосовують у виробництві ксиліту[16], ферментів, харчових добавок, для очищення від нафтових забруднень.
Також дріжджі широко використовують у науці як модельні організми для досліджень у генетиці, молекулярній і клітинній біології. Пекарські дріжджі (Saccharomyces cerevisiae) були першими з еукаріотів, у яких була повністю визначена послідовність геному ДНК.
Хлібопечення

Приготування печеного дріжджового хліба — одна з якнайдавніших технологій. У цьому процесі використовується переважно Saccharomyces cerevisiae. Вони проводять спиртне бродіння з утворенням безлічі вторинних метаболітів, які зумовлюють смакові і ароматичні якості хліба. Спирт випаровується при випічці. Крім того, у тісті формуються бульбашки вуглекислого газу, що змушують його «підніматися» і після випічки додають хлібові губчасту структуру і м'якість. Аналогічний ефект спричинає внесення до тіста соди і кислоти (зазвичай лимонної), але у цьому випадку не утворюються смакові сполуки.
Борошно зазвичай бідне цукрами, необхідними для бродіння, тому до тіста додають яйця або цукор. Для отримання більшої кількості смакових сполук тісто проколюють або перемішують, вивільняючи вуглекислий газ, а потім знову залишають «підніматися». З'являється, проте, ризик, що дріжджам не вистачить зброджуваного субстрату.
Виноробство

Дріжджі у природних умовах є на поверхні плодів, зокрема винограду, часто вони наявні у великій кількості у світлому восковому нальоті на ягодах, кутикулі. Особливо часто тут трапляється вид Hanseniaspora uvarum, хоча кутикула може містити і багато інших видів. Хоча «дикі» епіфітні дріжджі і можуть привести до непередбачуваного результату бродіння, зазвичай вони не витримують конкуренції з дріжджами, що мешкають у винних бочках.
Зібраний виноград давлять, отримуючи сік (муст, виноградне сусло) з 10—25 % цукру. Для отримання білих вин від нього відокремлюють суміш кісточок і шкірки (вичавки), у мусті для червоних вин вона залишається. Потім, у результаті бродіння, цукор перетворюється на етанол. Вторинні метаболіти дріжджів, а також сполуки, отримані з них при дозріванні вина, визначають його аромат і смак. Для отримання низки вин (наприклад шампанського) повторно зброджують вино, що вже перебродило.
Припинення бродіння пов'язане або з вичерпанням запасів цукрів (сухе вино), або з досягненням порогу токсичності етанолу для дріжджів. Хересні дріжджі, на відміну від звичайних дріжджів (які гинуть, коли концентрація спирту в розчині досягає 12 %), стійкіші. Спочатку хересні дріжджі були відомі тільки на півдні Іспанії (у Андалусії), де завдяки їхнім властивостям отримували міцне вино — херес (до 24 % за тривалої витримки). З часом хересні дріжджі були також виявлені у Вірменії, Грузії, Криму та ін. Хересні дріжджі використовують також у виробництві деяких міцних сортів пива.
Пивоваріння і квасоваріння

У пивоварінні як сировина використовується зерно (найчастіше ячмінь), що містить багато крохмалю, але мало зброджуваних дріжджами цукрів. Тому перед бродінням крохмаль гідролізують. Для цього використовуються амілази, утворені самим зерном при проростанні. Пророщений ячмінь носить назву солод. Солод розмелюють, змішують з водою і варять, отримуючи сусло, яке згодом зброджується дріжджами. Розрізняють пивні дріжджі низового і верхового бродіння (цю класифікацію ввів данець Хрістіан Хансен).
Дріжджі верхового бродіння (найпоширеніші — Saccharomyces cerevisiae) формують «шапку» на поверхні сусла, віддають перевагу температурам 14—25 °C (тому верхове бродіння також називаються теплим) і витримують вищі концентрації спирту. Дріжджі низового (холодного) бродіння (Saccharomyces uvarum, Saccharomyces carlsbergensis) мають оптимум розвитку за температури 6—10 °C і осідають на дно ферментера. У створенні пшеничного пива часто використовується Torulaspora delbrueckii. У виготовленні ламбіка застосовують дріжджі, що випадково потрапили у ферментер, зазвичай вони належать до роду Brettanomyces.
Квас виробляється за аналогічною схемою, проте, крім ячмінного, широко застосовують житній солод. До нього додають борошно і цукор, після чого суміш заливають водою і варять з утворенням сусла. Найважливішою відмінністю квасоваріння від виробництва пива є використання у зброджуванні сусла, крім дріжджів, молочнокислих бактерій.
Чайний гриб

Чайний гриб є симбіонтом кількох видів дріжджів та бактерій, що росте у солодкій рідині (зазвичай чай або сік), та зброджує її з отриманням кисло-солодкого напою — «чайного квасу». Вперше він згадується у китайських рукописах приблизно 250 року до н. е. Його використання у Російській імперії, зокрема в Україні, почалося у 1900-ті роки.
Чайний гриб є асоціацією дріжджів і оцтовокислих бактерій. Найчастіше спостерігаються асоціації дріжджів Brettanomyces bruxellensis, Candida stellata, Schizosaccharomyces pombe, Torulaspora delbrueckii, Zygosaccharomyces bailii та деяких інших, з рядом видів бактерій родини Acetobacteraceae[17].
У 50-ті роки 20 ст. в СРСР активно досліджували різні природні речовини для їхнього медичного використання, у процесі яких були відмічені антимікробні та протиатеросклеротичні властивості як зооглеї чайного гриба, так і рідини, в якій він вирощується[18].
Промислове виробництво спирту
Спиртове бродіння — процес, що зумволює утворення етанолу (CH3CH2OH) з водних розчинів вуглеводів (цукрів) під дією деяких видів дріжджів (див. ферментація) у процесі їхнього метаболізму.
У промисловості для виробництва спирту використовують цукрову тростину, фуражну кукурудзу та інші дешеві джерела вуглеводів. Для отримання зброджуваних моно- і олігосахаридів їх руйнують сірчаною кислотою або амілазами грибного походження. Потім проводиться зброджування і перегонка ректифікації спирту до стандартної концентрації близько 96 об. %[19]. Дріжджі роду Saccharomyces були генетично модифіковані для зброджування ксилози[20] — однієї з основних складових целюлози, що дозволило використовувати як сировину для отримання біоетанолу папір і відходи деревообробки. Все це знижує ціну та поліпшує становище біоетанолу в конкурентній боротьбі з вуглеводневим паливом[21].
Харчові та кормові дріжджі
Дріжджі багаті білками, їхній вміст може сягати 66 %, при цьому 10 % маси припадає на незамінні амінокислоти. Дріжджова біомаса може бути отримана з відходів сільського господарства, гидролізатів деревини, її вихід мало залежить від кліматичних і погодних умов. Тому її використання надзвичайно вигідне для збагачення білками їжі людини і корму сільськогосподарських тварин. Додавання дріжджів у ковбаси почалося ще у 1910-ті роки у Німеччині, у 1930-ті кормові дріжджі почали виробляти в СРСР, де ця галузь особливо розвинулася.

У СРСР перший великий завод з виробництва білка — папріну, потужністю 70 000 т на рік, був запущений 1973 року. Як сировина використовувалися відходи нафтопереробки. У 1980-ті роки в СРСР вироблялося 1 млн т дріжджового білка, що становило 2/3 від загальносвітових обсягів. Серед лідерів біотехнологічного виробництва дріжджового кормового білка і ліпофільно-жирових комплексів були НДР і Угорщина. Проте у 1990-ті роки у зв'язку з виниклими гігієнічними проблемами виробництва і застосуванням мікробного білка, а також з економічною кризою виробництво різко скоротилося. Дані, що накопичилися, свідчили про прояв ряду негативних ефектів від застосування папріну в годуванні птахів і тварин. З екологічних і гігієнічних причин знизився і інтерес до цієї галузі в усьому світі.
Проте на Заході зараз виробляють і продають різні дріжджові екстракти: вегеміт, марміт, бовріл, ценовіс[22]. Для отримання екстрактів використовуються або автолізати дріжджів (клітини руйнуються і білок стає доступним завдяки ферментам самих клітин), або їхні гідролізати (руйнування спеціальними речовинами). Вони застосовуються як харчові добавки і для додання стравам смакових якостей; крім того, існують косметичні засоби на основі дріжджових екстрактів.
Продають також дезактивовані (вбиті тепловою обробкою), але не зруйновані харчові дріжджі, особливо популярні у веганів через високий вміст білка і вітамінів (особливо групи B), а також малу кількість жирів. Деякі з них збагачені вітаміном B12 бактеріалього походження.
Застосування у медицині
- Висушені пивні дріжджі використовують для виробництва ліків і біологічно активних добавок.
- Тривалий час випускався препарат Гефефітін як загальнозміцнюючий лікарський засіб.
- Рідкі пивні дріжджі традиційно прописували ослабленим особам з алергічними захворюваннями і т. д.
- Існує ряд препаратів на основі Saccharomyces boulardii, що підтримують і поновлюють флору шлунково-кишкового тракту (пробіотики). Доведено, що S. boulardii знімає симптоми гострої діареї у дітей[23][24], знижує ризик виникнення різних видів діареї у дорослих[25][26][27], запобігає реінфекції Clostridium difficile[28], знижує частоту скорочень мускулатури кишечника у хворих на синдром подразненого кишечника[29].
Біоремедіація
Деякі дріжджі знаходять потенційне застосування у сфері біоремедіації (біологічного очищення території). Одним з прикладів є дріжджі Yarrowia lipolytica, які перероблюють пальмову олію у сточних водах млинів[30], ТНТ (вибуховий матеріал)[31] та багато інших вуглеводнів, наприклад алкани, жирні кислоти, жири й олії[32].
Дослідження як модельного організму
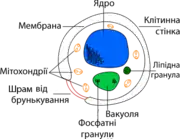
Кілька видів дріжджів, особливо Saccharomyces cerevisiae, широко використовуються у генетиці і клітинній біології як модельний організм. Це значною мірою відбувається тому, що клітинний цикл та фізіологічні процеси клітин дріжджів дуже подібні до відповідних процесів людських клітин, і тому основні клітинні механізми реплікація ДНК, рекомбінація, поділ клітини і метаболізм мають багато спільних рис[33]. Також багато білків, важливих у біології людини, вперше були знайдені при вивченні їхніх гомологів у дріжджах; ці білки включають білки клітинного циклу, сигнальні білки і ферменти, що модифікують білки.
24 квітня 1996 року було заявлено, що S. cerevisiae став першим еукаріотом, чий геном, що складається з 12 млн пар основ, був повністю секвенований у рамках геномного проєкту[12]. У той час це був найскладніший організм, чий геном був секвенований, цей процес зайняв 7 років і залучав понад 100 лабораторій[34]. Другим видом дріжджів, чий геном був секвенований, був Schizosaccharomyces pombe, завершений у 2002 році[35]. Це був шостий геном еукаріотів розміром 13,2 млн пар основ.
Дріжджі як фактор псування продуктів
Дріжджі здатні рости на середовищах з низьким pH (5,5 і навіть нижче), особливо у присутності вуглеводів, органічних кислот і інших легко утилізованих джерел органічного вуглецю[36]. Вони добре розвиваються за температур 5—10°С, коли міцеліальні гриби вже нездатні до росту.
У процесі життєдіяльності дріжджі метаболізують компоненти харчових продуктів, утворюючи власні специфічні кінцеві продукти метаболізму. При цьому фізичні, хімічні і, як наслідок, органолептичні властивості продуктів змінюються — продукт «псується»[37]. Розростання дріжджів на продуктах нерідко видно неозброєним оком як поверхневий наліт (наприклад на сирі або на м'ясних продуктах) або проявляють себе, запускаючи процес бродіння (у соках, сиропах і навіть у достатньо рідкому варенні).
Дріжджі роду Zygosaccharomyces вже тривалий час є одними з найважливіших агентів псування продукції харчової промисловості. Особливо ускладнює боротьбу з ними той факт, що вони можуть рости за високих концентрацій цукрози, етанолу, оцтової кислоти, бензойної кислоти і диоксиду сірки[38], що є найважливішими консервантами.
Патогенні дріжджі
Деякі види дріжджів є факультативними і умовними патогенами, зазвичай спричиняючи захворювання у людей з ослабленою імунною системою.
Дріжджі роду Candida є компонентами нормальної мікрофлори людини, проте у разі загального ослаблення організму травмами, опіками, хірургічним втручанням, тривалого застосування антибіотиків, у ранньому дитячому віці і в старості дріжджі цього роду можуть масово розвиватися, спричиняючи захворювання — кандидоз. Існують різні штами цього гриба, зокрема достатньо небезпечні. У нормальних умовах у людському організмі дріжджі роду Candida обмежуються у своєму розвитку природною бактеріальною мікрофлорою людини (лактобактерії та ін.), але у випадку розвитку патологічного процесу багато з них утворюють високопатогенні співтовариства з бактеріями.
Ще один вид дріжджів, Cryptococcus neoformans, викликає криптококкоз, особливо небезпечний для ВІЛ-інфікованних людей: серед них захворюваність криптококкозом сягає 7—8 % у США і 3—6 % у Західній Європі. Клітини C. neoformans оточені міцною полісахаридною капсулою, яка перешкоджає розпізнаванню і знищенню їх лейкоцитами. Дріжджі цього виду найчастіше виявляють у посліді птахів, причому самі птахи не хворіють.
Рід Malassezia включає в себе облігатних симбіонтів теплокровних тварин і людини, що не трапляються ніде, окрім їхніх шкірних покривів. При порушеннях імунітету вони викликають пітиріаз (строкатий лишай), фолікуліт і себорейний дерматит. У здорових людей за нормального функціонування сальних залоз Malassezia ніяк себе не проявляє і навіть відіграє позитивну роль, перешкоджаючи розвиткові небезпечніших патогенів.
Примітки
- Hakim, Ahmad (23 травня 2019). Yeast FAQ's. Phaff Yeast Culture Collection (англ.). Процитовано 7 лютого 2022.
- Gamero, Amparo; Quintilla, Raquel; Groenewald, Marizeth; Alkema, Wynand; Boekhout, Teun; Hazelwood, Lucie (1 грудня 2016). High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiology (англ.) 60. с. 147–159. ISSN 0740-0020. doi:10.1016/j.fm.2016.07.006. Процитовано 7 лютого 2022.
- Kurtzman, C.P., Fell, J.W. (2006). Yeast Systematics and Phylogeny — Implications of Molecular Identification Methods for Studies in Ecology. Biodiversity and Ecophysiology of Yeasts, The Yeast Handbook, Springer. Архів оригіналу за 22 лютого 2008. Процитовано 7 січня 2007.
- Bass D.; Howe, A.; Brown, N.; Barton, H.; Demidova, M.; Michelle, H.; Li, L.; Sanders, H.; Watkinson, S.C.; Willcock, S.; Richards, T.A. (16 жовтня 2007). Yeast forms dominate fungal diversity in the deep oceans. Proc Biol Sci. ISSN 0962-8452.
- Kurtzman, C.P., Fell, J.W. 2006. Yeast systematics and phylogeny — implications of molecular identification methods for studies in ecology. In: Rosa, C.A. and Peter, G., editors. The Yeast Handbook. Germany: Springer-Verlag Berlin Herdelberg. p. 11-30.
- Walker K, Skelton H, Smith K. (2002). Cutaneous lesions showing giant yeast forms of Blastomyces dermatitidis. J Cutan Pathol. 29 (10): 616–618. PMID 12453301.
- Ostergaard, S., Olsson, L., Nielsen, J. (2000). Metabolic Engineering of Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 64: 34–50.
- Бабьева И. П., Чернов И. Ю. (2004). Биология дрожжей. Москва: Товарищество научных изданий КМК.
- Yeast, The Contemporry Review (1871), Collected Essays VIII..
- Planets in а Bottle, More about Yeast Архівовано 4 листопада 2009 у Wayback Machine., Science@NASA
- Barnett, James A. (2003). Beginnings of microbiology and biochemistry: the contribution of yeast research. Microbiology 149: 557—567. PMID 12634325. Архів оригіналу за 28 квітня 2007. Процитовано 26 серпня 2007.
- Williams, N (April 26, 1996). Genome Projects: Yeast Genome Sequence Ferments New Research. Science 272 (5261): 481–0. doi:10.1126/science.272.5261.481.
- COMPLETE DNA SEQUENCE OF YEAST
- Schizosaccharomyces pombe: Second yeast genome sequenced. Архів оригіналу за 3 травня 2008. Процитовано 26 серпня 2007.
- Balasubramanian M., Bi E., Glotzer M. (2004). Comparative analysis of cytokinesis in budding yeast, fission yeast and animal cells. Curr Biol. 14 (18): R806–R818. PMID 15380095.
- R. Sreenivas Rao, R.S. Prakasham, K. Krishna Prasad, S. Rajesham, P.N. Sarma, L. Venkateswar Rao (2004) Xylitol production by Candida sp.: parameter optimization using Taguchi approach, Process Biochemistry 39:951-956
- «Yeast ecology of Kombucha fermentation» Архівовано 11 жовтня 2007 у Wayback Machine.. International Journal of Food Microbiology.
- «Чайный гриб и его лечебные свойства» (Г. Ф. Барбанчик, 1954)
- «Fuel Ethanol Production». Genomics: GTL.
- «Genetically Engineered Saccharomyces Yeast Capable of Effective Cofermentation of Glucose and Xylose». American Society for Microbiology.
- «Yeast rises to a new occasion» Архівовано 30 серпня 2007 у Wayback Machine.. American Society for Microbiology.
- Тулякова Т. В., Пасхин А. У., Седов В. Ю. Дрожжевые екстракти — безпечні джерела вітамінів, мінеральних речовин і амінокислот // Харчова промисловість № 6, 2004. Архівовано 8 жовтня 2007 у Wayback Machine.
- Centina-Sauri G, Sierra Basto G (1994). Therapeutic evaluation of Saccharomyces boulardii in children with acute diarrhea. Ann Pediatr 41: 397–400.
- Kurugol Z, Koturoglu G (2005 Jan). Effects of Saccharomyces boulardii in children with acute diarrhoea. Acta Paediatrica 94: 44–47.
- McFarland L, Surawicz C, Greenberg R (1995). Prevention of β-lactam associated diarrhea by Saccharomyces boulardii compared with placebo. Am J Gastroenterol 90: 439–48.
- Kollaritsch H, Kemsner P, Wiedermann G, Scheiner O (1989). Prevention of traveller's diarrhoea. Comparison of different non-antibiotic preparations. Travel Med Int: 9–17.
- Saint-Marc T, Blehaut H, Musial C, Touraine J (1995). AIDS related diarrhea: a double-blind trial of Saccharomyces boulardii. Sem Hôsp Paris 71: 735–41.
- McFarland L, Surawicz C, Greenberg R (1994). A randomised placebo-controlled trial of Saccharomyces boulardii in combination with standard antibiotics for Clostridium difficile disease. J Am Med Assoc 271: 1913–8.
- Maupas J, Champemont P, Delforge M (1983). Treatment of irritable bowel syndrome with Saccharomyces boulardii: а double blind, placebo controlled study. Medicine Chirurgie Digestives. 12(1): 77–9.
- Oswal, N; Sarma PM, Zinjarde SS, Pant A. (Oct 2002). Palm oil mill effluent treatment by a tropical marine yeast.. Bioresour Technol. 85 (1). PMID 12146640.
- Jain, MR; Zinjarde SS, Deobagkar DD, Deobagkar DN (Nov 2004). 2,4,6-trinitrotoluene transformation by a tropical marine yeast, Yarrowia lipolytica NCIM 3589.. Mar Pollut Bull. 49 (9-10): 783–8. PMID 15530522.
- Fickers, P; Benetti PH, Wache Y, Marty A, Mauersberger S, Smit MS, Nicaud JM (April 2005). Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications.. FEMS Yeast Res. 5 (6-7): 527–543. PMID 15780653.
- «What are yeasts?» Архівовано 26 лютого 2009 у Wayback Machine.. Saccharomyces Genome Database. Отримано 24 грудня 2006.
- COMPLETE DNA SEQUENCE OF YEAST. Отримано 21 січня 2007.
- Schizosaccharomyces pombe: Second yeast genome sequenced. Отримано 21 січня 2007.
- Kurtzman, C.P. 2006. Detection, identification and enumeration methods for spoilage yeasts. In: Blackburn, C. de. W, editor. Food spoilage microorganisms. Cambridge, England: Woodhead Publishing. р. 28-54.
- Fleet, G.H., and Praphailong, W., Yeasts, In: Spoilage of Processed Foods: Causes and Diagnosis, AIFST (2001), Southwood Press. p 383–397
- Fugelsang, K.C., Zygosaccharomyces, A Spoilage Yeast Isolated from Wine Архівовано 1 січня 2008 у Wayback Machine., California Agriculture Technology Institute.
Посилання
- Дріжджі // Універсальний словник-енциклопедія. — 4-те вид. — К. : Тека, 2006.
- Yeast growth and the cell cycle
- Yeast virtual library
- Science of Breadmaking
- Ancient Egyptian Bread Making
- Brewing Yeast Propagation and Maintenance: Principles and Practices
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |