Плазмодесма
Плазмоде́сма — цитоплазматичний канал вистелений плазматичною мембраною, що пронизує клітинні стінки сусідніх рослинних клітин, сполучаючи їх між собою. Основною функцією плазмодесм є забезпечення спілкування і транспорту між клітинами, що інакше були б ізольовані одна від одної мертвими клітинними стінками. Через те, що майже всі рослинні клітини з'єднані таким чином, їхні цитоплазми складають один неперервний симпласт. Внутрішньоклітинний транспорт через плазмодесми називається симпластним[1].
| Плазмодесма | |
| |
Функціональними аналогами плазмодесм у тварин є щілинні контакти[2].
Схожі структури, які спостерігали в мікроскоп у ціанобактерій називали мікроплазмодесмами.[3]
Утворення плазмодесм
У залежності від походження розрізняють первинні та вторинні плазмодесми. Первинні плазмодесми формуються під час процесу цитокінезу. По завершенню мітозу між двома новоутвореними клітинами починає утворюватись клітинна пластинка шляхом злиття везикул апарату Гольджі, які містять попередники речовин клітинної стінки. Подекуди злиття везикул порушується наявністю залишків веретена поділу та трубочок гладкого ендоплазматичного ретикулуму, в цих місцях цитоплазми клітин залишаються сполученими. По мірі відкладання додаткових шарів полімерів клітинної стінки утворені канали видовжуються і перетворюються в плазмодесми. Таким чином первинні плазмодесми забезпечують зв'язок між клонально спорідненими клітинами[1].
Вторинні плазмодесми утворюються у вже сформованих клітинних стінках. Вони можуть виникати шляхом випинання плазматичної мембрани або галуженням наявних плазмодесм. Вторинні плазмодесми можуть сполучати як клонально споріднені так і не споріднені клітини[1].
Будова плазмодесм
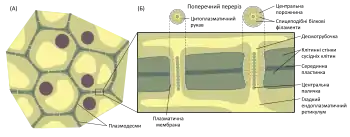
Діаметр плазмодесм коливається від 20 до 50 нм, довжина визначається сумою товщини клітинних стінок сусідніх клітин та серединної пластинки, типово становить щонайменше 0,1 мкм[2]. Плазмодесми мають складну внутрішню будову, потрібну для регуляції транспорту макромолекул між клітинами[1].
В центрі більшості плазмодесм розташована тоненька циліндрична трубочка (десмотрубочка), що сполучає порожнини гладкого ендоплазматичного ретикулуму сусідніх клітин. Зовнішня сторона мембрани десмотрубочки та внутрішня сторона мембрани, що вистилає плазмодесму, вкриті глобулярними білками. Ці білки сполучаються між собою спицеподібними філаментами, які поділяють цитоплазматичний рукав на 8—10 мікроканалів.
Дослідження руху флуоресцентних барвників через плазмодесми дозволило встановити їхню пропускну межу (англ. size exclusion limit, SEL). З'ясувалось, що найбільші молекули, здатні транспортуватись через цитоплазматичний рукав, мають молекулярну масу 700—1000 да, що відповідає розміру приблизно 1,5—2 нм. Проте ця величина може регулюватись[1].
Регулювання транспорту через плазмодесми
Плазмодесми є динамічними структурами, вони можуть не тільки утворюватись і зникати, а й змінювати свої властивості, зокрема SEL, в залежності від умов. У деяких експриментах із флуоресцентними барвниками було показано, що транспорт молекул значно менших за 700 да між певними типами клітин може бути заблокований, не зважаючи на те, що вони сполучені морфологічно нормальними плазмодесмами[2]. З іншого боку, були отриманні електронні мікрофотографії, що демонстрували проходження через цитоплазматичний рукав вірусних частинок діаметром 10 нм. Вважається, що віруси рослин мають спеціальні білки руху (англ. viral movement proteins), які зв'язуються із плазмодесмами і розширюють їх, уможливлюючи транспорт вірусної РНК або цілих вірусних частинок[4].
Вірусні білки руху імітують дію фізіологічних систем рослини, що забезпечують збільшення SEL плазмодесм. Ці системи є особливо активними під час розвитку рослинного організму, в цей час між групами клітин у пагоні та корені повинен відбуватись транспорт регуляторних білків, що визначають шлях диференціації. Ці білки зв'язуються із плазмодесмами і збільшують їхню пропускну здатність, після чого проникають у сусідню клітину. Таким чином також транспортуються деякі молекули мРНК[2].
Мікроплазмодесми
Мікроплазмодесми — тонкі пори, розташовані в перегородках (септах) певних нитчастих бактерій (деяких ціанобактерій та актиноміцетів), що з'єднують сусідні клітини в трихоми (нитці). Ці пори використовуються для комунікації та обміну метаболітами між двома сусідніми клітинами, які відокремлені перегородкою.
Джерела
- Tiaz L., Zeiger E. (2010). Plant Physiology (вид. 5th). Sinauer. ISBN 978-0-87893-507-9. Архів оригіналу за 9 грудня 2012. Процитовано 6 березня 2012.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Wilk, Laura; Strauss, Mike; Rudolf, Mareike; Nicolaisen, Kerstin; Flores, Enrique; Kühlbrandt, Werner; Schleiff, Enrico (2011). Outer membrane continuity and septosome formation between vegetative cells in the filaments of Anabaena sp. PCC 7120. Cellular Microbiology 13 (11): 1744–1754. ISSN 14625814. doi:10.1111/j.1462-5822.2011.01655.x.
- Campbell NA, Reece JB (2008). Biology (вид. 8th). Benjamin Cammings. ISBN 978-0321543257.
Рекомендована література
Gunning B. E. S., Robards A. W. (Eds.): Intercellular Communications in Plants: Studies on Plasmodesmata, Springer-Verlag, New York, 1976 - ISBN 978-3-642-66296-6
