Щілинні контакти
Щілинні конта́кти або нексуси — це тип міжклітинних контактів, що характеризуються наявністю білкових каналів, які дозволяють проходження іонів та невеликих молекул між сусідніми клітинами, таким чином зв'язуючи їх метаболічно та електрично (інколи терміном «щільний контакт» позначають самі канали[1]). У ділянці контакту цього типу між мембранами залишається щілина шириною 2—4 нм, звідки і походить їхня назва. Проте найпринциповішим елементом нексусу є не сама щілина, а канали з білків конексинів або інексинів, що регулярно її перетинають[2][3].
Щілинні контакти були вперше виявлені у збудливих тканинах, таких як м'язова та нервова, де формують електричні синапси, згодом було з'ясовано, що вони існують і в епітеліальній та сполучній тканинах багатоклітинних тварин[2].
Щілинні контакти за біологічним значенням аналогічні до плазодесм рослин із тією відмінністю, що останні можуть пропускати макромолекули, такі як білки та РНК[2].
Будова і молекулярний склад
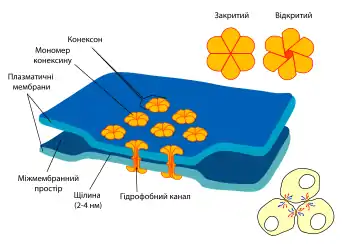
Щілинні контакти мають форму круглих зон діаметром 0,5—5 мкм[4], у яких зосереджено від кількох до кількох тисяч міжклітинних каналів. Ці канали складаються у безхребетних складаються із білків інексинів (12 генів у Caenorhabditis elegans і 25 у Drosophila melanogaster[2]), а в хребетних переважно із конексинів (21 ген в людини, позначаються Cx[2]) — білків, що виникли внаслідок конвергентної еволюції[5], хоча в геномах хребетних збереглись і споріднені з інексинами гени панексинів. Конексини та інексини хоча й схожі за формою і функціями, але не мають гомолічних амінокислотних послідовностей[2].
Конексини — це білки із молекулярною масою 25—50 кДа[3], що перетинають мембрану чотири рази[2][5]. Шість таких молекул формують гексагональний циліндр — напівканал або конексон, який має діаметр приблизно 7—8 нм і просвіт шириною в 1,5 нм[4]. Два конексони у мембранах сусідніх клітин з'єднуються між собою кінець в кінець формуюючи повний канал. Щілинний контакт складається із великої кількості таких каналів. Оскільки конексони утримують мембрани на певній відстані одна від одної між ними зберігається щілина сталої ширини (2—4 нм)[4].
Щілинні контакти різних тканин відрізняються властивостями через те, що вони побудовані із різних комбінацій конексинів. Більшість клітин експресують більше одного гену конексинів, продукти яких можуть сполучатись між собою утворюючи гомомерні, тобто такі що складаються із однакових субодиниць, або гетеромерні конексони, проте дозволена тільки обмежена кількість комбінацій. Крім того конексон може контактувати на сусідній клітині із ідентичним або відмінним за білковим складом напівканалом[2][5].
Динаміка і регуляція
Щільні контакти є динамічними структурами і можуть швидко утворюватись, руйнуватись та змінюватись. Дослідження із флуоресцентно міченими конексинами показали, що нові конексони постійно додаються на периферії щілинного контакту, а старі забираються із середини та руйнуються. Час півжиття одного конексона становить кілька годин. Нові конексони додаються шляхом злиття мембранних везикул із плазмалемою, після чого вони мігрують в її площині поки не досягнуть периферії нексуса і не сполучаться із конексонами на сусідній клітині. Таким чином у плазматичні мембрані постійно існує невелика кількість неспарених конексонів, які за нормальних умов підтримуються у закритій конформації[2][5].
Проникність конексонів може регулюватись, наприклад у збудливих тканинах вони часто є потенціал-керованими. Канали щільних контактів швидко закриваються під впливом збільшення цитозольного pH. До схожих наслідків призводить із підвищення концентрації кальцію, це пояснюється тим, що за нормальних умов в цитоплазмі клітини підтримується низький рівень цього іону. При ушкодженні клітини її мембрана може «протікати», внаслідок чого всередину проникають катіони Na+ та Ca2+, а назовні — цінні метаболіти. Якщо б клітина за таких умов залишалась з'єднаною із сусідніми, то вони б також зазнали негативних змін[3][2].
На проникність щілинних контактів в нервовій тканині також можуть впливати нейромедіатори, наприклад у сітківці ока вона знижується при дії дофаміну[5].
Функції

Одні із перших даних про щілинні контакти були отримані в електрофізіологічних дослідженнях, в яких було виявлено, що електричне збудження може швидко і безпосередньо передаватись між нейронами певного типу. Для визначення діаметра пор щілинних контактів в одну із клітин, сполучених таким чином, молекули різного розміру мічені флуоресцентними барвниками та спостерігали їх проникнення у сусідні клітини. Таким чином було встановлено, що максимальний діаметр пори становить близько 1,5 нм, а отже вона може пропускати неорганічні іони, моносахариди, амінокислоти, нуклеотиди, вітаміни та вторинні посередники, такі як цАМФ, інозитолтрифосфат тощо, але не макромолекули[2].
Щілинні контаки є важливими для передачі сигналів між електрично збудливими клітинами. У нервовій системі вони формують електричні синапси, які дозволяють значно швидше поширення потенціалу дії ніж хімічні. Такі структури вигідні у випадках, коли швидкість відіграє ключову роль, наприклад у деяких рефлекторних дугах, що відповідають за втечу, у риб і комах, а також, коли група нейронів повинна діяти синхронно. У хребетних тварин щілинні контакти сполучають клітини серцевого м'яза, синхронізуючи їхні скорочення, а також клітини гладеньких м'язів кишківника, забезпечуючи їхні ритмічні перестальтичні скорочення[2].
У незбудливих тканинах щілинні контакти забезпечують вирівнювання концентрації сигнальних молекул та поживних речовин. Наприклад, вони є необхідними для нормального функціонування печінки, зокрема її відповіді на сигнали від нервових терміналій, що контактують лише із частиною гепатоцитів. Також щілинні контакти потрібні для дозрівання фолікулів у яєчнику: вони сполучають ооцит та гранульозні клітини[2]. Вони забезпечують явище так званої метаболічної кооперації (метаболічного спряження), необхідної для супресії соматичних мутацій. Наприклад при синдромі Леша-Ніхана, який виникає внаслідок порушення функцій ферменту гіпоксантинфосфорибозилтрансферази, клітини нездатні утворювати один із нуклеотидів, потрібних для синтезу ДНК — дАТФ. Якщо такі клітини культивувати разом із здоровими, то вони будуть отримувати достатню кількість цього метаболіту через щілинні контакти. Вважається що саме через наявність щілинних контактів гетерозиготні по синдрому Леша-Ніхана жінки мають нормальний фенотип. Оскільки ген гіпоксантинфосфорибозилтрансферази розташований в X-хромосомі, випадкова інактивація однієї із двох копій цих хромосом мала би призводити до виникнення мозаїки нормальних та мутантних клітин[5][3].
Також щілинні контакти потрібні для нормального розвитку зародка. У хребетних більшість клітин на ранніх стадіях ембріогенезу електрично спряжені. В процесі диференціації окремих груп клітин вони від'єднуються від інших, проте залишаються сполученими між собою. Таким чином кожна група може поводитись синхронно і відмінно від інших[2].
Хвороби пов'язані із щілинними контактами
Зважаючи на різноманітність функцій щілинних контактів в організмі людини, очевидно, що мутації в білках, які входять до їх складу, можуть стати причиною широкого спектра порушень від катаракти (мутації у Cx46 або Cx50) до демієлінізації периферичних нервів (мутації у Cx32). Наприклад мутації у генах багатьох конексинів, особливо Cx26, є найчастішими причинами спадкової глухоти, що виникає внаслідок загибелі клітин кортієвого органа; мутації у гені конексину, експресія якого відбувається у фолікулах, призводить до безпліддя[2].
Джерела
- Harvey Lodish et al. (2000). [http://www.ncbi.nlm.nih.gov/books/NBK21607/def-item/A7517/ Glossary]. Molecular Cell Biology (вид. 4th). W H Freeman. ISBN 0-7167-3136-3. — Переглянуто 26 березня 2012
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Harvey Lodish et al. (2000). [http://www.ncbi.nlm.nih.gov/books/NBK21502/ 15.7 Transport across Epithelia]. Molecular Cell Biology (вид. 4th). W H Freeman. ISBN 0-7167-3136-3. — Переглянуто 26 березня 2012
- Луцик О. Д., Іванова А. Й., Кабак К. С., Чайковський Ю. Б. (2003). Гістологія людини. Київ: Книга плюс. ISBN 966-7619-39-7.
- Goodenough DA, Paul DL (2009). Gap junctions. Cold Spring Harb Perspect Biol 1. PMID 20066080. doi:10.1101/cshperspect.a002576.
