Репарація ДНК
Репарація ДНК (від англ. DNA repair — «ремонт ДНК») — набір процесів, за допомогою яких клітина знаходить і виправляє пошкодження молекул ДНК, які кодують її геном. У клітинах всіх організмів, зокрема людини, як нормальна метаболічна активність, так і зовнішні фактори, такі як ультрафіолетове випромінювання, можуть викликати пошкодження ДНК, приводячи у людини до 1 мільйона індивідуальних молекулярних пошкоджень на клітину за день[1]. Багато з цих пошкоджень заподіює структурні пошкодження молекулі ДНК і може впливати на транскрипцію генів клітини, або навіть повністю запобігати їй. Деякі пошкодження викликають потенційно шкідливі мутації в геномі клітин, які впливають на виживання клітини або її дочірніх клітин після мітозу. Тому процеси репарації ДНК повинні бути постійно активними, оскільки вони мають швидко відповідати на будь-які пошкодження структури ДНК.
Ефективність репарації ДНК залежить від багатьох факторів, зокрема типу клітини, її віку та оточення. Клітини, що накопичили велику кількість пошкоджень ДНК або не мають більше можливостей до репарації, можуть піти одним з трьох можливих шляхів: перейти до незворотного статичного стану або клітинного старіння, піддатися апоптозу або альтруїстичному самогубству, або продовжити ділитися і стати першою клітиною ракової пухлини.
Репарація ДНК життєво важлива для підтримки цілісності геному і тому для нормального функціонування організму. Багато генів, які, як було показано, впливають на тривалість життя, пізніше виявилися залученими в репарацію і захист ДНК[2]. Нездатність виправлення деяких молекулярних пошкоджень генеративних клітин можуть ввести мутації в геном нащадків і таким чином впливають на швидкість еволюції.
Порушення ДНК бувають декілька різних типів. Це можуть бути одноланцюгові, дволанцюгові розриви молекули ДНК, піримідинові димери, модифікації нуклеотидів, найчастішими з яких є алкілування азотистих основ, метилювання чи етилування O- та N-атомів нуклеотидів. В залежності від типу порушень механізми їх репарації відрізняються[3].[4]
Ексцизійні репарації
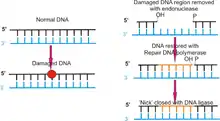
Під час ексцизійної репарації (від лат. excisio «вирізати») молекула ДНК сканується, знаходяться пошкоджені ділянки і видаляється нуклеотид або ціла частина одного ланцюгу ДНК навколо пошкодженого нуклеотиду. Вилучене місце «залатується» новосинтезованими нуклеотидами, які добудовуються комплементарно іншому, неушкодженому ланцюгу ДНК.
Ексцизійна репарація буває[4]:
- Ексцизійна репарація азотистих основ (BER), видаляє і заміняє один нуклеотид.
- Ексцизійна репарація нуклеотидів (NER), видаляє і заміняє ділянку одного ланцюга ДНК. У бактерій E.coli ексцизійну репарацію після мутагенного впливу ультрафіолету виконує комплекс urvABC (англ. ultra violet repair)[3]
Некомплементарні пари основ (місметч)
Репарація некомплементарних пар основ дозволяє знайти ділянку ДНК, де на різних ланцюгах навпроти один одного сформувалась некомплементарна пара нуклеотидів (наприклад, А-Ц замість А-Т). Це відбувається за допомогою комплексу mutHLSU з чотирьох білків: mutS розпізнає місметч, mutL формує структуру, на яку сідає два білки mutH, що є нуклеазою, а mutU, геліказа, розплітає ДНК. Далі ДНК-полімераза повинна добудувати видалені нуклеотиди. Цікаво, що видаляється новосинтезований ланцюг ДНК і розпізнавання відбувається завдяки тому, що в складі старого, метеринського ланцюга ДНК є метильовані цитозини, які відсутні в тільки-що створеному дочірньому.
Дволанцюгові розриви
Дволанцюгові розриви молекули ДНК репаруються за допомогою консервативного процесу негомологічного з'єднання кінців. У ссавців більшість дволанцюгових розривів саме так і репаруються. При наявності більше одного подвійного розриву репарація може призвести до хромосомних аберацій. Також у генній інженерії використовують систему CRISPR/Cas9 для спеціального, цільового мутагенезу і вирізання частини гену шляхом двох двуланцюгових розривів.
Одразу після реплікації ДНК, а також у деяких організмів, можливе точне з'єднання порушення ДНК за допомогою гомологічної рекомбінації[4].
У кожної хромосоми є кінці — теломери, які могли бути розпізнані як дволанцюговий розрив ДНК. Але спеціальна структура теломер запобігає цьому. Але при кожному поділі відбувається скорочення теломер і врешті-решт вони вкорочуються до такої межі, що не можуть формувати специфічну структуру — це так звана межа Гейфліка, і з нею пов'язана одна з теорій старіння.
При постреплікативній репарації реплісома може обійти пошкодження, залишивши прогалину, але на її місце шляхом гомологічної рекомбінації вставляється ділянка сестринської молекули ДНК.
Примітки
- Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. (2004). Molecular Biology of the Cell, p963. WH Freeman: New York, NY. 5th ed.
- Browner WS, Kahn AJ, Ziv E, Reiner AP, Oshima J, Cawthon RM, Hsueh WC, Cummings SR. (2004). The genetics of human longevity. Am J Med 117(11):851-60. DOI:10.1016/j.amjmed.2004.06.033
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет". с. 300–323.
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика. К: Видавничо-поліграфічний центр "Київський університет". с. 35–46.
Посилання
- Репарація ДНК[недоступне посилання з липня 2019] — Біологія людини(рос.)
- Репарація ДНК — www.cellbiol.ru(рос.)
Джерела
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет". с. 300–323.
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика. К: Видавничо-поліграфічний центр "Київський університет". с. 35–46.
