CAM-фотосинтез
CAM фотосинтез (англ. Crassulacean acid metabolism) — спосіб фіксації вуглецю, який розвивався на деяких рослин як пристосування до посушливих умов.[1] У рослині, що використовує повний САМ-фотосинтез, продих в листі залишається закритим протягом дня, щоб зменшити евапотранспірацію, але відкритий вночі для збору вуглекислого газу (CO2). CO2 зберігається як малат з чотирьох вуглецевих кислот у вакуолях вночі, а потім у денний час малат переносять до хлоропластів, де він повертається до CO2, який потім використовується під час фотосинтезу. Заздалегідь зібраний CO2 концентрується навколо ферменту RuBisCO, що підвищує ефективність фотосинтезу. Цей метаболізм спочатку вивчався у рослин родини Crassulaceae. Вони в основному включають сукуленти. Вперше, коли вивчали CAM-фотосинтез, Crassula використовувався як модельний організм.

Історія
CAM-фотосинтез спочатку запідозрив де Соссюр у 1804 р. У його Recherches Chimiques sur la Vegetation, підтверджений та удосконалене Е.Обертом в 1892 р. У своїй книзі «Фізиологічні дослідження лікарських рослин» та викладене Річардсом, 1915 р. В «Кислотному та газовому обміні» в Какті, Інститут Карнегі. Термін ''CAM-фотосинтез'', можливо, був винайдений ботаніками Рансоном і Томасом у 1940 році, але вони не були першими, хто виявив цей цикл. Це спостерігали ботаніки Ренсон і Томас, у соковитій сім'ї Crassulaceae (що включає нефритні рослини і Sedum)[2]. Її назва відноситься до метаболізму кисню в Crassulaceae, а не про метаболізм «крейсулолової кислоти»
Огляд: дві частини циклу
CAM-фотосинтез є адаптацією для підвищення ефективності використання води, і, як правило, вона зустрічається в рослинах, що виростають в посушливих умовах.[3]
Протягом ночі
Рослина, що використовує CAM-фотосинтез, має свої відкриті місця, дозволяючи вводити CO2 і бути фіксованими як органічні кислоти, які зберігаються у вакуолях. Протягом дня продихи закриваються (таким чином, запобігаючи втраті води), і вуглець виділяється до циклу Кальвіна, щоб фотосинтез мав місце.
Двоокис вуглецю фіксується в цитоплазмі клітин мезофілу за допомогою реакції з PEP, подібної до реакції С4. Але, на відміну від механізму С4, отримані органічні кислоти зберігаються у вакуолях для подальшого використання; Тобто вони не відразу надходять до циклу Кальвіна. Останній не може працювати протягом ночі, тому що легкі реакції, які забезпечують його АТФ і НАДФН, не можуть відбутися.
Впродовж дня
Протягом дня органічні кислоти, що зберігають CO2, виділяються з вакуолей мезофілових клітин і надходять у строму хлоропластів, де фермент вивільняє CO2, який потім входить в цикл Кальвіна.
Переваги
Найважливішою перевагою CAM-фотосинтезу для рослин є здатність залишати більшість листових продихів закритими протягом дня. Рослини, що використовують цей тип фотосинтезу, найчастіше зустрічаються в західних умовах, де вода припадає на преміум. Здатність тримати продих закритою протягом найгарячішої і найсухішої частини дня зменшує втрату води через евапотранспірацію, що дозволяє таким рослинам рости в середовищах, які в іншому випадку були б занадто сухими. Рослини, що використовують лише фіксацію вуглецю C3, наприклад, втрачають 97 % води, яку вони поглинають через коріння до транспірації, — це висока вартість, яку уникають рослини, здатні використовувати CAM-фотосинтез.
Порівняння з метаболізмом С4
C4 шлях має схожість з CAM; обидва діють, щоб концентрувати CO2 навколо RuBisCO, тим самим підвищуючи його ефективність. CAM-фотосинтез концентрує його тимчасово, забезпечуючи CO2 протягом дня, а не вночі, коли дихання є домінуючою реакцією. C4 рослини, навпаки, концентрують CO2 просторово, з реакційним центром RuBisCO в «клітинах провідного пучка», заповненої CO2. Внаслідок бездіяльності, необхідної механізмом CAM, C4 вуглецева фіксація має більшу ефективність в плані синтезу 3-фосфогліцеринової кислоти.
Біохімія
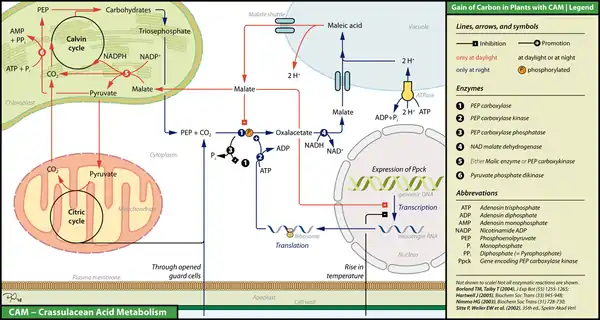
Рослини з CAM повинні контролювати зберігання CO2 та його зниження до розгалужених вуглеводів у просторі та часі.
При низьких температурах (часто вночі) рослини, що використовують CAM, відкривають свої продихи, молекули CO2 дифундують у внутрішньоклітинних просторів губчатого мезофілу, а потім у цитоплазму. Тут вони можуть зустріти фосфоенолпіруват (PEP), який є фосфорильованим триозою. За цей час рослини синтезують протеїн, що називається PEP-карбоксилазакіназа (PEP-C кіназа), експресія якого може бути інгібована високими температурами (часто при денному світлі) та наявністю малату. PEP-C кіназа фосфорилює його цільовий фермент PEP-карбоксилаза (PEP-C). Фосфорилювання різко підвищує здатність ферменту каталізувати утворення оксалоацетату, який потім може бути перетворений на малат за допомогою малатдегідрогенази NAD +. Малат потім транспортується через матальні човники у вакуолі, де його перетворюють у форму зберігання ялинової кислоти. На відміну від PEP-C-кінази, PEP-С синтезується весь час, але майже пригнічується при денному світлі або дефосфорилювання через PEP-C фосфатазу, або безпосередньо шляхом зв'язування малату. Останнє неможливе при низьких температурах, оскільки малат ефективно транспортується у вакуолі, тоді як ПЕП-С кіназа легко інвертує дефосфорилювання.
Під час денного світла рослини, що використовують САМ, закривають свої захисні клітини та виділяють малат, який потім транспортується в хлоропласти. Там, залежно від виду рослин, він розщеплюється на піруват та CO2 яєчним ферментом або PEP карбоксикіназою. CO2 потім вводять у цикл Кальвіна, сполучену і саморегенераційну ферментну систему, яка використовується для створення розгалужених вуглеводів. Побічний продукт пірувату може бути додатково деградований у циклі цитохмії мітохондріальної кислоти, тим самим забезпечуючи додаткові молекули CO2 для циклу Кальвіна. Піруват може також використовуватися для відновлення PEP через піруват-фосфат дикіназу, етап високої енергії, який вимагає АТФ та додаткового фосфату. Під час наступної прохолодної ночі PEP, нарешті, експортується в цитоплазму, де він бере участь у фіксації вуглекислого газу через малат.
Див. також
Джерела
- C.Michael Hogan. 2011. Respiration. Encyclopedia of Earth. Eds. Mark McGinley & C.J.cleveland. National council for Science and the Environment. Washington DC
- Ranson S. L.; Thomas M (1960). «Crassulacean acid metabolism». Annual Review of Plant Physiology.
- Herrera, A. (2008), «Crassulacean acid metabolism and fitness under water deficit stress: if not for carbon gain, what is facultative CAM good for?», Annals of Botany, 103 (4): 645—653
