Фотодихання
Фотодихання (гліколатний шлях, C2-фотосинтез) — стимульоване світлом виділення вуглекислого газу та поглинання кисню в рослин переважно з C3-типом фотосинтезу. Також під фотодиханням розуміють біохімічний шлях пов'язаний з регенерацією однієї молекули 3-фосфогліцеринової кислоти (C3) із двох молекул гліколевої кислоти (C2), який лежить в основі описаного вище газообміну. Наявність біохімічного механізму фотодихання обумовлене значною оксигеназною активністю РуБісКО, ключового ферменту циклу Кальвіна.
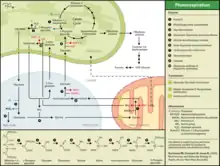
Поглинання кисню в ході фотодихання обумовлено оксигеназною активністю РуБісКО в хлоропластах і роботою оксидази гліколевої кислоти в пероксисомах. Крім того окиснення утвореного в мітохондріях НАДН також пов'язане з поглинанням кисню. Виділення вуглекислого газу (C1) при фотодиханні відбувається в мітохондріях і пов'язане з конденсацією двох молекул гліцину (C2) з утворенням однієї молекули серину (C3) (послідовна робота двох ферментів: гліцин декарбоксилази та серингідроксиметилтрансферази). Також в реакції конденсації гліцину в мітохондріях виділяється аміак, який реутилізується в результаті роботи глутамінсинтетази та глутаміноксоглутаратамінотрансферази (ГС/ГОГАТ шлях). При фотодиханні витрачається АТФ (не відбувається запасання енергії) синтезований в ході фотофосфорилювання. Також окиснення гліколевої кислоти в пероксисомах в ході фотодихання є основним джерелом токсичного пероксиду водню у рослинної клітини, яка фотосинтезує.
Історія
Перші свідчення фотодихання були отримані 1920 року німецьким біохіміком Отто Варбургом[1]. При дослідженні водоростей роду Chlorella було показано інгібування фотосинтезу (поглинання вуглекислого газу) при підвищенні концентрації кисню. Цей ефект спостерігався як при високих, так і при низьких інтенсивностях світла та пізніше отримав назву ефекту Варбурга[2][3].
Газообмін і відмінності від темнового дихання
Сумарне поглинання кисню на світлі визначається інтенсивністю двох процесів: темнового дихання та світлоіндукованого дихання, пов'язаного з фотохімічними реакціями хлоропластів. Фотодихання активується при високих інтенсивностях світла, в той час як процеси темнового дихання рослин подавляються на світлі. Фотодихання збільшується при підвищенні концентрації кисню від 0 до 100 %, а темнове дихання насичується вже при 2 % кисню[4][5]. На відміну від темнового дихання процес поглинання кисню при фотодиханні не інгібується типовими дихальними отрутами, наприклад азид натрію (інгібітор цитохромоксидази мітохондрій).
Субстратна специфічність РуБісКО

Рибулозобісфосфаткарбоксилаза/оксигеназа (РуБісКО) — ключовий фермент фотосинтезу, каталізує дві конкуруючі реакції: карбоксилювання та оксигенування п'ятивуглецевого цукру рибулозо-1,5-бісфосфату (РуБФ). Як і при карбоксилюванні молекула кисню приєднується до ендіольної форми РуБФ зв'язаної з ферментом, яка існує внаслідок кето-енольної ізомерії. Незважаючи на те, що константа Міхаеліса (концентрація напівнасичення) для вуглекислого газу набагато нижча (9 мкМ), ніж для кисню (535 мкМ), тобто спорідненість ферменту до вуглекислого газу значно вища[6]. Швидкість оксигеназної реакції висока, оскільки концентрація кисню в атмосфері становить 21 %, а вуглекислого газу 0,035 — 0,038 %. Також дається взнаки більша розчинність кисню у воді порівняно з вуглекислим газом. Завдяки цьому у C3-рослин інтенсивність фотодихання може досягати 50 % від інтенсивності фотосинтезу.
Хімізм фотодихання
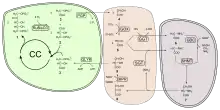
Здійснення реакцій фотодихання потребує тісної взаємодії трьох органел рослинної клітини: хлоропластів, пероксисом і мітохондрій. Через наявність оксигеназної активності у ферменту рибулозобісфосфаткарбоксилази, що каталізує приєднання CO2 до рибулозо-1,5-біфосфату на початковій стадії циклу Кальвіна, замість асиміляції вуглекислоти під дією того ж ферменту можливе окиснення рибулозофосфату з розпадом його на 3-фосфогліцеринову кислоту, яка може надійти у цикл, і на фосфат гліколевої кислоти. Він дефосфорилюється і гліколева кислота транспортується в пероксисоми, де окиснюється до гліоксалевої кислоти і амінується з отриманням гліцину. В мітохондріях із двох молекул гліцину утворюється серин і вуглекислий газ. Серин може використовуватися для синтезу білка, або перетворюватися в 3-фосфогліцеринову кислоту та повертається у цикл Кальвіна.
Умови, що стимулюють фотодихання
Очевидно, що зниження концентрації вуглекислого газу призводить до стимуляції фотодихання. Як було сказано вище, фотодихання інтенсифікується і при збільшенні концентрації кисню. Збільшення температури призводить до зниження стійкості ендіольного інтермедіату реакції, що каталізується РуБісКО, що сприяє реакції оксигенування РуБФ. Крім того, при підвищенні температури розчинність газів у воді зменшується, що призводить до зниження концентрації вуглекислого газу, в той час як концентрація кисню змінюється мало.
Біологічне значення фотодихання
Хоча достовірно відомо, що фотодихання знижує ефективність фотосинтезу, призводить до втрат асимільованого вуглецю, тим не менше питання про функції фотодихання залишається дискусійним. Основна гіпотеза припускає, що фотодихання виникло як шлях, що слугує для найоптимальнішої утілизації гліколату, який утворюється в результаті оксигеназної активності РуБісКО. При цьому той факт, що оксигеназна активність РуБісКО не була елімінована в ході еволюції пояснюється, мабуть тим, що співвідношення карбоксилазної та оксигеназної активностей досягло межі, що визначається хімізмом реакції та не може бути збільшене. Порівняльний аналіз ферменту різних організмів показує, що РуБісКО існувала вже 3,5 млрд років тому, коли в атмосфері було мало кисню і вже до того часу зайняла ключову позицію в циклі асиміляції вуглецю при фотосинтезі. При цьому її оксигеназна функція в умовах низького вмісту кисню не відігравала суттєвої ролі. При збільшенні вмісту кисню втрати асимільованого вуглецю в результаті фотодихання зростали, однак складність будови РуБісКО, мабуть, завадила еволюції каталітичного центру для усунення оксигеназної активності[6]. Ця гіпотеза опосередковано підтверджується відсутністю значних успіхів у спробах генноінженерним способом внести зміни в амінокислотну послідовність активного центру РуБісКО, які збільшили б спорідненість ферменту до вуглекислого газу[7].
У зв'язку з цим в ході еволюції в ряду рослин виникли механізми, що мінімізують фотодихання, не пов'язані з модифікацією РуБісКО. До таких механізмів належать різні типи C4-фотосинтезу та CAM-фотосинтез. У цих біохімічних шляхах первинну фіксацію вуглекислоти здійснює фосфоенолпіруваткарбоксилаза (ФЕП-карбоксилаза), що дозволяє в результаті концентрувати вуглекислоту в місці її асиміляції в реакції карбоксилювання РуБФ, яка каталізується РуБісКО.
Див. також
Примітки
- Чиков В. И. Фотодыхание // Соросовский образовательный журнал, 1996, № 11, c. 2-8. Архів оригіналу за 18 серпня 2007. Процитовано 23 лютого 2017.
- Turner JS, Brettain EG (February 1962). Oxygen as a factor in photosynthesis. Biol Rev Camb Philos Soc 37: 130–70. PMID 13923215. doi:10.1111/j.1469-185X.1962.tb01607.x.[недоступне посилання з травня 2019]
- Zelitch I (1971). Chapter 8, Section E: Inhibition by O2 (The Warburg Effect). Photosynthesis, Photorespiration, and Plant Productivity. New York: Academic Press. с. 253–255. ISBN 0124316085.
- Физиология растений / под ред. И. П. Ермакова. — М. : «Академия», 2007. — 640 с. — ISBN 978-5-7695-36-88-5.
- Фотосинтез. Физиолого-экологические и биохимические аспекты / А.Т Мокроносов, В. Ф. Гавриленко, Т. В. Жигалова; под ред. И. П. Ермакова. — М. : «Академия», 2006. — 448 с. — ISBN 5-7695-2757-9
- Биохимия растений / Г.-В. Хелдт; пер. с англ. — М. : БИНОМ. Лаборатория знаний, 2011. — 471 с. — ISBN 978-5-94774-795-9
- Spreitzer RJ, Salvucci ME (2002). Rubisco: structure, regulatory interactions, and possibilities for a better enzyme. Annu Rev Plant Biol 53: 449–75. PMID 12221984. doi:10.1146/annurev.arplant.53.100301.135233.
Література
- Физиология растений / под ред. И. П. Ермакова. — М. : «Академия», 2007. — 640 с. — ISBN 978-5-7695-36-88-5. (рос.)
- Фотосинтез. Физиолого-экологические и биохимические аспекты / А.Т Мокроносов, В. Ф. Гавриленко, Т. В. Жигалова; под ред. И. П. Ермакова. — М. : «Академия», 2006. — 448 с. — ISBN 5-7695-2757-9 (рос.)
- Биохимия растений / Г.-В. Хелдт; пер. с англ. — М. : БИНОМ. Лаборатория знаний, 2011. — 471 с. — ISBN 978-5-94774-795-9 (рос.)
- Чиков В. И. Фотодыхание // Соросовский образовательный журнал, 1996, № 11, c. 2-8 (рос.)
- Plant Physiology, Fifth Edition — Sinauer Associates, Inc. Taiz and Zeiger. 2010. (англ.)
Посилання
- Фотодихання // Універсальний словник-енциклопедія. — 4-те вид. — К. : Тека, 2006.