Ceratioidei
Ceratioidei або глибоководні вудильники — підряд костистих риб ряду Вудильникоподібні (Lophiiformes). Підряд включає 11 родин та 120 видів. Цей підряд ззовні відрізняється від інших вудильникоподібних насамперед відсутністю черевних плавців. У той час як інші вудильникоподібні мешкають на дні або, в небагатьох випадках, біля поверхні, ці вудильники постійно живуть на великій глибині, мешкаючи у відкритому океані в товщі води. У цьому підряді проявляється статевий диморфізм, а в окремих родинах, і, мабуть, незалежно, виробився статевий паразитизм самців.
? Ceratioidei | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
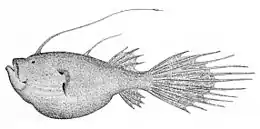 Ceratias holboelli | ||||||||||||||||||
| Біологічна класифікація | ||||||||||||||||||
| ||||||||||||||||||
Родини | ||||||||||||||||||
|
Дивитись текст | ||||||||||||||||||
Посилання | ||||||||||||||||||
| ||||||||||||||||||
Спосіб життя
Розмноження
Незважаючи на те що дорослі глибоководні вудильники живуть на глибинах, куди не проникає світло і де відсутні будь-які сезонні зміни, в Північній Атлантиці, де вони вивчені найбільше, всі види розмножуються у весняну або літню пору. Нерест відбувається, очевидно, на великих глибинах. Розвиваються ікринки поступово піднімаються догори, і личинки завдовжки 2-3 мм вилуплюються у приповерхневому шарі 30-200 м, де живляться переважно веслоногими рачками і планктонними щетинкощелепними (Chaetognatha). До початку метаморфозу молодь встигає опуститися на глибину понад 1000 м. Мабуть, їхнє занурення відбувається швидко, так як самиці на стадії метаморфоза зустрічаються в шарі 2-2,5 тис. м, а самці на цій же стадії — на глибині 2 тис. м. В шарі 1500–2000 м мешкають обидві статі, що пройшли метаморфоз і досягли статевої зрілості, але іноді дорослі особини зустрічаються і на менших глибинах. Дорослі самки живляться в основному глибоководними батипелагічними рибами (міктофові, хауліоди, топірцями, меламфаями та ін.) ракоподібними й рідше головоногими, а дорослі самці, подібно личинкам, — веслоногими рачками і щетинкощелепними. Пов'язані з індивідуальним розвитком вертикальні міграції глибоководних вудильників мають дуже велике пристосувальне значення, так як тільки у приповерхневому шарі їх малорухливі і численні личинки можуть знайти достатньо корму, щоб накопичити запаси для майбутнього метаморфоза. Величезні втрати внаслідок поїдання ікри і личинок хижаками компенсуються у вудильників дуже великою плодючістю. Їхня ікра дрібна (не більше 0,5 — 0,7 мм в діаметрі). Їхні прозорі личинки нагадують крихітні балончики, завдяки тому що вони одягнені в шкіряний чохол, роздутий студенистою тканиною. Ця тканина збільшує плавучість і розміри личинок, що поряд з прозорістю оберігає їх насамперед від дрібних хижаків, найбільш небезпечних для них своєю численністю.
Статевий диморфізм
Всі глибоководні вудильники характеризуються дуже різко вираженим статевим диморфізмом. Уже на самих ранніх личинкових стадіях, коли за формою тіла, розмірами, ступенем розвитку очей і нюхових органів обидві статі аж до стадії метаморфозу подібні між собой, самки відрізняються від самців наявністю «вудки», в яку перетворений змінений перший промінь спинного плавця.
.png.webp)
Під час і особливо після метаморфозу у самиць відносна величина голови і рота сильно збільшується, нюхові органи далі не розвиваються, очі, як правило, стають маленькими, а у найбільших видів майже зовсім дегенеруються. У самців, навпаки, тіло стає більш видовженим, відносна величина голови і щелеп сильно зменшується, очі залишаються великими, а нюхові органи досягають дуже великих розмірів.
У самиць, що пройшли метаморфоз, на щелепах, леміші і верхньоглоткових кістках розвиваються тонкі гострі зуби, своїми загнутими вершинами спрямовані назад і нерідко здатні складатися всередину. У багатьох видів вони дрібні і численні, але у деяких щелепні зуби дуже довгі (Linophryne, Lasiognathus та ін.). У тауманіхта (Thaumanichthys pagidostomus) вони особливо сильно розвинені на верхній щелепі, а у неоцерації (Neoceratias spinifer) з'являються додаткові величезні зуби, що сидять прямо на рилі і зовні нижньої щелепи.
У самців, що пройшли метаморфоз, замість личинкових зубів в передній частині рота розвиваються особливі зуби, не пов'язані з щелепами і зливаються своїми основами. Вони діють як щипчики для захоплення дрібної здобичі і для прикріплення до самиць.
Мабуть, найяскравіше статевий диморфізм проявляється в розмірах. Все вільноживучі самці досягаютьзавдожки не більше 16-46 мм. Самиці набагато більші. Хоча у більшості видів вони не перевищують завдовжки 5-10 см, у декількох видів самиці досягають 20 см, у криптопсара (Cryptopsaras couesi) до 44 см, а у гімантолофа (Himantolophus groenlandicus) — до 60 см і ваги 4 кг. Найбільший самець у гімантолофа має довжину всього 46 мм і вагу 0,82 г, тобото по довжині він менший за самицю у 13 раз, а за вагою у 5000 разів. Однак найбільшими за всіх є самиці церації (Ceratias holboelli), що досягають завдовжки понад 1 м.
Статевий паразитизм
У чотирьох родинах глибоководних вудильників (Caulophrynidae, Ceratiidae, Neoceratiidae і Linophrynidae) існують абсолютно незвичайні відносини між статями, що не зустрічаються серед інших риб і невідомі для хребетних тварин. Вони виражаються в тому, що карликові самці живуть у вигляді паразитів на тілі самок. Самець прикріплюється в шкіру самки гострими щипчикоподібними зубами. Незабаром губи і язик такого самця повністю зливаються з тілом самки, а його щелепи, зуби, очі і кишечник поступово редукуються так, що врешті-решт він перетворюється на придаток, що виробляє сперму. Живлення паразитуючого самця здійснюється за рахунок крові самки, так як її кровоносні судини з'єднуються з судинами самця. Але зябра самця і крихітні зяброві отвори зберігаються, внаслідок чого кров, що надходить до нього може збагачуватися киснем. Відмінності в розмірах самки і паразитуючого на ній самця можуть бути величезними. Так, наприклад, у самки церації (Ceratias holboelli) завдовжки 119 см і вагою 7 кг прикріплений до неї самець мав довжину 16 мм і важив всього 14 мг. У решти 7 видів, у яких виявлений паразитизм самців ( Саіlophryne jordani, Neoceratias spinifer, Cryptopsaras couesi, Edriolychnus schmidti, Photocorinus spiniceps, Borophryne apogon і Linophryne argyresca), після прикріплення до самок самці, мабуть, вже не збільшуються в розмірах і не перевищують в довжину 14-22 мм.
Паразитуючі самці прикріплюються тільки до дорослих самкам і лише після того, як вони самі завершать стадію метаморфози. На одній і тій же самці на різних ділянках її тіла може одночасно прикріплюватися до трьох самців. Одного разу прикріпившись, такі самці назавжди втрачають самостійність і, судячи з усього, протягом ряду років беруть участь в розмноженні. До переходу до паразитизму такі самці володіють добре розвиненими очима і дуже великими нюховими органами. Це дозволяє припускати, що вони відшукують самок за специфічним запахом, сліди якого зберігаються довгий час в практично нерухомій воді великих глибин. Наблизившись до самки, самець, очевидно, може візуально «уточнити» її приналежність до свого виду за будовою вудки, освітлюваної спалахами її світного органу, або ж за кольором і частотю самих спалахів, закодованих відповідним чином. Самці інших шести родин глибоководних вудильників, мабуть, ведуть вільний спосіб життя, але не виключена ймовірність, що й вони в період нересту тимчасово прикріплюються до самок.
Опис


Тільки у самок глибоководних вудильників зберігається «вудилище» (ілліцій) з «приманкою» (ескою), яка світиться, що у різних видів сильно варіює за формою і величиною і буває забезпечена найрізноманітнішими шкірними придатками. У самок більшості видів ілліцій короткий, але в особин деяких родів(Rhynchactis, Lasiognathus і Gigantactis) він дуже довгий. Так, у гігантактіса (Gigantactis macronema) ілліцій в 4 рази перевищує довжину тіла. У лазіогната (Lasiognathus saccostoma) базальна частина ілліція має вигляд довгого прута, що втягується в спеціальну піхву, а його тонка і гнучка кінцева частина вінчається ескою з трьома гачками. Вся це споруда виглядає як справжня оснащена вудка. Настільки ж незвичайну будову має ілліцій у церації (Ceratias holboelli): базальна частина його дуже сильно подовжена і розташовується в спеціальному каналі на спині, де вона може вільно висуватися або втягуватися. Підманюючи здобич, цей вудильник поступово присуває «приманку» (еску) до величезного рота і в потрібний момент заковтує жертву. У своєріднії придонної галатеатауми (Galatheathauma axeli), з глибини близько 3600 м «приманка» розташована в роті. На відміну від інших глибоководних вудильників галатеатаума полює, мабуть, не в товщі води, а лежачи на дні.
Світний орган на есці являє собою залозу, заповнену слизом, в якій живуть бактерії, що світяться. Завдяки розширенню стінок артерій, що постачають кров'ю цю залозу, риба довільно може викликати світіння бактерій, які потребують для цього припливу кисню, або припиняти його, звужуючи відповідні судини. Зазвичай світіння відбувається у вигляді серії послідовних спалахів. У самок деяких родин (Diceratiidae і Сеratiidae) є додатковий булавоподібний світний орган на другому головному промені, причому у самок церацій, крім того, перед м'яким спинним плавником є 1-3 подібних органи. У родині лінофринових у самок є особливі придатки на підборідді. Можливо, що у лінофріни (Linophryne arborifera) величезний деревовидний підбородковий придаток теж несе світні залози.
Дорослі вудильники забарвлені в темно-коричневий або чорний колір, їх тіло зазвичай голе, і луска є лише в окремих видів. Так, у тауманіхта (Thaumanichthys pagidostomus) шипики маються на черевній стороні тіла, а у гімантолофа (Himantolophus) на тілі є кісткові бляшки, часом дуже великі.

Шлунок самок глибоководних вудильників здатний дуже сильно розтягуватися, завдяки чому вони можуть заковтувати і перетравлювати дуже велику здобич, нерідко перевершуючу їх по своїх розмірах. Жадібність вудильників часом призводить до загибелі не тільки їхньої жертви, але і їх самих. Іноді на поверхні знаходять мертвих вудильників із заковтнутою рибою, що перевищує їх за розмірами більш ніж в два рази. Захопивши таку велику здобич, вудильник не може її випустити завдяки будові своїх зубів, і йому нічого не залишається, як продовжувати заковтувати рибу, яка прагне звільнитися з капкана. До того часу, коли вудильник закінчує обід, він гине, мабуть, від втрати сил.
Поширення
Личинки глибоководних вудильників зустрічаються лише в тропічній і помірно теплій зонах Світового океану, що лежать між 40° пн.ш. і 35° пд.ш. і обмежених літніми ізотермами 20°С в поверхневих водах. У більш високих широтах, включаючи субарктичні і субантарктичні води, зустрічаються тільки дорослі особини, які потрапляють туди завдяки винесенню їх течіями.
Родини
- Підряд Ceratioidei
- Centrophrynidae
- Ceratiidae
- Himantolophidae
- Diceratiidae
- Melanocetidae
- Thaumatichthyidae
- Oneirodidae
- Caulophrynidae
- Neoceratiidae
- Gigantactinidae
- Linophrynidae
Посилання
- Kurt Fiedler: Lehrbuch der Speziellen Zoologie, Band II, Teil 2: Fische, Gustav Fischer Verlag, Jena 1991, ISBN 3-334-00339-6
- Joseph S. Nelson: Fishes of the World, John Wiley & Sons, 2006, ISBN 0-471-25031-7
- E. O. Wiley, G. David Johnson: A teleost classification based on monophyletic groups. in Joseph S. Nelson, Hans-Peter Schultze, Mark V. H. Wilson: Origin and Phylogenetic Interrelationships of Teleosts. Verlag Dr. Friedrich Pfeil, München 2010, ISBN 978-3-89937-107-9
- Theodore W. Pietsch & Christopher P. Kenaley: Ceratioidei. Seadevils, Devilfishes, Deep-sea Anglerfishes, 2007 (The Tree of Life Web Project)
