Інтрон
Інтро́н — ділянка ДНК, яка є частиною гену, але на відміну від екзонів, не містить інформації про послідовність амінокислот білка.[1][2]
Послідовність нуклеотидів, що відповідає інтрону, видаляється з транскрибованої з нього РНК в процесі сплайсингу до того, як відбудеться трансляція (зчитування білка). Інтрони характерні для всіх типів еукаріотичної РНК, але також знайдені в невеликій кількості в рибосомальній РНК (рРНК) і транспортній РНК (тРНК) прокаріотів. Число і довжина інтронів дуже різні в різних видах і серед різних генів одного організму. Наприклад, геном риби фугу (Takifugu rubripes) містить мало інтронов порівняно з іншими хребетними тваринами. З другого боку, гени ссавців і квіткових рослин часто містять численні інтрони, які можуть бути значно довше екзонів.
Загальні відомості
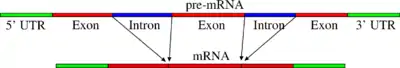
Хоча інтрони і не несуть інформації про послідовність амінокислот у білку, вони іноді дозволяють альтернативний сплайсинг гену, таким чином, що розділені інтроном ділянки можуть комбінуватися у різній послідовності, виробляючи різні варіанти білка з одного гену. Контроль сплайсингу мРНК і, в результаті, вибір кінцевого продукту, виконується широкою різноманітністю сигнальних молекул. Інтрони також іноді містять «застарілий код» секції гену, що ймовірно колись був частиною білка, але потім перестав використовуватися.
Загалом вважається, що послідовність інтронів є ділянкою ДНК без певної функції. Проте зараз це піддається сумніву: відомо, що інтрони містять декілька коротких послідовностей, які важливі для ефективного сплайсингу. Точний механізм дії цих інтронних енхансерів ще недостатньо відомий, але вважається, що вони служать обов'язковими ділянками ефективного синтезу білків, що стабілізують сплайсосому. Також можливо, що вторинна структура РНК, сформована з використанням послідовності інтронів, також має ефект на сплайсинг, інтронна послідовність при одному альтернативному сплайсингу стає екзонною при іншому. Послідовності «застарілого коду», з другого боку, в більшості випадків дійсно є «еволюційними артефактами».
Перший опис інтрону було зроблено в 1977 році Філіпом Шарпом і Річардом Робертсоном незалежно один від одного, за що вони були нагороджені Нобелівською премією з фізіології і медицини в 1993 року. Термін «інтрон» ввів американський біохімік Волтер Гілберт (англ. Walter Gilbert) в 1978 році.
Деякі інтрони, наприклад інтрони групи I і II, фактично є рибозимами, які здатні до каталізу свойого власного сплайсингу із первинної копії РНК. За це відкриття Чех Томас отримав Нобелівську премію з хімії, яку він розділив у 1989 році з Сіднеєм Алтманом. Крім того, інтрони можуть брати участь в експресії генів.
Класифікація інтронів
Зараз класифіковані чотири групи інтронів:
- Ядерні інтрони
- Інтрони групи I
- Інтрони групи II
- Інтрони групи III
Іноді інтрони групи III також відносять до групи II, тому що вони схожі по струкурі і функціям. Ядерні або спалайсосомні інтрони піддаються сплайсингу при допомозі сплайсосоми і малих ядерних РНК (snoRNA). У послідовності РНК, що містить ядерні інтрони, є спеціальні сигнальні послідовності, які пізнаються сплайсосомою.
Інтрони I, II і III груп здатні до автосплайсингу і зустрічаються рідше, ніж сплайсосомні інтрони. Інтрони II і III групи схожі один на одного і мають збережену вторинну структуру. Вони володіють властивостями, схожими на властивості сплайсосоми і, ймовірно, є її еволюційними попередниками. Інтрони I групи, які трапляються у бактерій і найпростіших, — єдиний клас інтронів, що вимагає присутності незв'язаного гуанідинового нуклеотиду. Їхня вторинна структура відрізняється від вторинної структури інтронів II і III групи.
Еволюція інтронів
Існують дві альтернативні теорії, що пояснюють походження й еволюцію сплайсосомних інтронів. Це так звані теорії ранніх інтронів (РІ) і пізніх інтронів (ПІ). Теорія РІ передбачає, що численні інтрони були присутні в загальних предках представників всіх доменів життя і, відповідно, є дуже старими структурами. Згідно з цією моделлю, інтрони були практично втрачені з геному прокаріотів. Також вона передбачає, що ранні інтрони сприяли рекомбінації екзонів, що представляють домени білків. ПІ пропонує, що інтрони з'явилися в генах відносно недавно, і були інсертовані (вставлені) в геном після розділення організмів на окремі домени. Ця модель ґрунтується на спостереженні, що сплайсосомні інтрони зустрічаються тільки у еукаріотів.
Ідентификація
Майже всі еукаріотичні ядерні інтрони починаються з GU й закінчуються на AG (правило AG-GU).