Сплайсосома
Сплайсосома — структура, що складається з молекул РНК і білків і що здійснює видалення некодуючих послідовностей (інтронів) з попередників мРНК під час процесингу РНК. Цей процес називається сплайсингом (від англ. splicing — зрощення, стикування). Сплайсосому складають п'ять малих ядерних рибонуклеопротеїнів (мяРНП, у їх склад входять мяРНК) і деяка кількість додаткових білкових факторів.
Ці процеси здійснюються мяРНП, що містяться в сплайсосомі, які називаються U1, U2, U4, U5 і U6, та більш ніж 100 білками, які не асоційовані з процесом сплайсингу. Вони беруть участь в багатьох взаємодіях між молекулами РНК, а також між РНК і білками. РНК-частина мяРНП багата урацилом.
У загальному випадку збірка сплайсосоми відбувається заново для кожного мРНК-попередника (пре-мРНК). Молекула пре-мРНК обов'язково містить специфічні фрагменти послідовності, що розпізнаються під час збірки сплайсосоми. Це 5'-кінець, послідовність точки відгалуження, поліпирімідинова ділянка і 3'-кінець.
Сплайсосома каталізує видалення проміжних послідовностей і з'єднання сусідніх екзонів. Інтрони зазвичай визначаються по наявності послідовності GU на 5'-кінці і послідовностей AG на 3'-кінці. 3'-кінець може бути також визначений по ланцюжку поліпіримідинів різної довжини, званої поліпіримідиновим трактом (ППТ). ППТ виконують функцію скріплення факторів з 3'-кінцем, а також, можливо, зв'язування факторів з послідовністю точки відгалуження (ПТВ). ПТВ, у свою чергу, містить багато аденіну, потрібного для початку сплайсинга.
Група менш поширених мяРНП — U11, U12, U4atac і U6atac — входить разом з U5 в склад малої сплайсосоми, що виконує сплайсинг рідкісних інтронів пре-мРНК, званих інтронами типу U12.
Дослідження сплайсосоми
Ранні дослідження, в першу чергу порівняння послідовностей кДНК і геномних клонів, показали, що інтрони були широко поширені у вищих еукаріотичних генах. Ці аналізи також виявили наявність консенсусних та консервативні послідовності, які створюють межі інтронів.
Проте, біохімічний аналіз сплайсингу довів існування неклітинних систем (екстрактів отриманих з клітин), які були здатні каталізувати ефективне і точне зшивання синтетичних пре-мРНК. Наявність таких систем сприяла з'ясуванню механізму сплайсингу і дозволила ідентифікувати клітинні компоненти, які є необхідними для цього процесу.
Безліч експериментів, в тому числі седиментаційний аналіз показав, що, при інкубації з екстрактом, премРНК утворювали комплекси, аналогічні за розміром до рибосом. Пізніше було встановлено, що ці структури були сплайсосомами, які здійснювали сплайсинг мРНК.
Відкриття сплайсосоми призвело до інтенсивного вивчення механізму цього процесу та встановлення структури цієї системи.
Сплайсинг РНК
Робота Шарпа і Робертса, опублікована в 1977 році, показала, що гени вищих організмів «розсіяні», тобто зберігаються в декількох різних ділянках ДНК[1][2]. Кодуючі відрізки гена перемежаються з некодуючою ДНК, яка не використовується при експресії генів. «Розсіяна» структура гена була виявлена, коли аденовірусна мРНК була схрещена з фрагментами моноспіралі ДНК, що залишилася після дії ендонуклеази[1]. Після такого схрещування з'ясувалося, що мРНК-ділянки цих гібридних молекул мРНК-ДНК містять 5'- і 3'-кінці ділянок, що не мали воднемих зв'язків. Довші відрізки ДНК при гібридизації закільцевувалися і утворювали відгалуження. Стало ясно, що ці закільцьовані ділянки, що містять непотрібні послідовності, витягуються з пре-мРНК в результаті процесу, який і був названий «сплайсингом». Згодом було також з'ясовано, що така розсіяна структура украй широко поширена серед еукаріотичних генів.
Альтернативний сплайсинг, під час якого відбувається рекомбінація екзонів — головне джерело генетичного різноманіття у еукаріотів. Змінному зрощенню зазвичай приписувалася роль причини малої кількості генів в геномі людини. Оцінка їх кількості впродовж багатьох років мінялася, максимальна ж кількість прогнозувалася на рівні 100 тисяч. Проте завдяки проекту геному людини, а також його подальшої анотації, стало відомо, що їх кількість може бути менше 19 тисяч.[3] Разом з тим, загальна кількість відомих білків людини за базою даних UniProt перевищує 154 тисячі станом на липень 2016 року.[4]
Збірка сплайсосоми
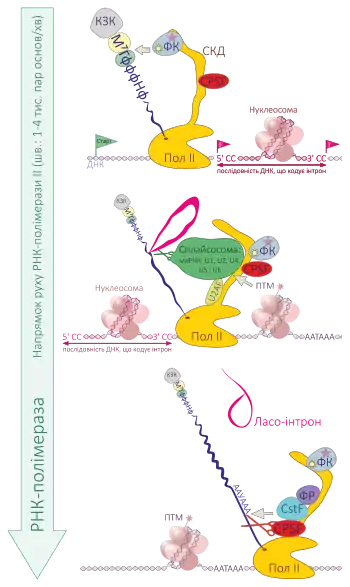
Канонічна модель формування активної області сплайсосоми є впорядкованим покроковим процесом з'єднання одиниць мяРНП на підкладці пре-мРНК. Під час первинного розпізнавання пре-мРНК відбувається зв'язування мярРНП U1 з 5'-стиком пре-мРНК, а також формування з інших білкових факторів фіксаційного комплексу або раннього комплексу (Е-комплексу) для ссавців[5][6]. Фіксаційний комплекс — АТФ-незалежна формація, що утримує пре-мРНК для здійснення сплайсингу[7].
Малий ядерний рибонуклеопротєїн U2 зв'язується з областю відгалуження за рахунок взаємодії з компонентом Е-комплексу U2AF (допоміжним фактором мяРНП U2) і, можливо, з U1. В результаті АТФ-залежної реакції U2 утворює міцний зв'язок з послідовністю точки відгалуження (ПТВ), тим самим формуючи комплекс А. Подвійний зв'язок між U2 і областю відгалуження пре-мРНК витягує аденін з ланцюжка, що відгалужується[8]. Присутність в U2 псевдоуридинових залишків практично напроти області відгалуження приводить до зміни конфігурації зв'язків РНК-РНК під час зв'язування з U2. Ці зміни структури, викликані псевдоуридином, поміщають 2-OH-групу витягнутого аденіну в положення, що дозволяє зробити перший крок сплайсингу[9].
U4, U5 і U6, що зв'язуються з утворенням більшого потрійного мяРНП, зв'язуються із сплайсосомою, що збирається, і формують комплекс В, який після деяких проміжних перегруповувань субодиниць перетворюється на комплекс C — власне сплайсосому, яка приступає до каталізу сплайсингу[10][11].
Неясно, яким чином потрійний мяРНП U4-U5-U6 зв'язується з комплексом А. Цей процес може бути опосередуваним взаємодією між білками, так само як і взаємодіями між нуклеотидами мяРНК, що входять до складу U2 і U6.
U5 взаємодіє з послідовностями на 5'- і 3'-кінцях сплайсингової ділянки за рахунок інваріантної петлі мяРНК, що входить до його складу[12]. Білкові компоненти U5 взаємодіють з 3-регіоном сплайсингової ділянки[13].
Після зв'язування з потрійним мяРНП і перед початком сплайсингу в сплайсосомі відбувається безліч змін конфігурації зв'язків РНК-РНК. Ці зміни продовжуються сплайсосомі, що вже почала роботу. Багато взаємодій між РНК є взаємовиключними, проте досі неясно, що викликає ці взаємодії і завдяки чому дотримується їх порядок.
Перше перетворення — це, можливо, зсув U1 з 5’-кінця сплайсингової ділянки і виникнення там зв'язку з U6. Відомо, що U1 слабо пов'язаний з тією, що діє сплайсосомою[14], а також є інгібітором для утворення зв'язку між U6 і 5-концом (показано на прикладі короткого ланцюжка РНК, 5-экзон, що містить, і 5-конец сплайсингової ділянки[15]).
Зв'язування U2 з ПТВ — ще один приклад взаємодії між РНК і білками. При зв'язуванні U2 із сплайсосомою, білок Е-комплексу SF1, що зв'язується з ділянкою відгалуження, витісняється, оскільки наявність U2 виключає його зв'язування з цією ділянкою. Усередині самого U2 також відбуваються деякі взаємовиключні зміни конфігурації. Наприклад, в його активній формі, виникає шпилька IIa, а неактивній же формі домінує взаємодія між шпилькою і подальшою ділянкою ланцюжка[11].
Неясно, за рахунок чого U4 відділяється від U6. Вважається, що в збірці сплайсосоми бере участь декілька геліказ, які можуть розкручувати РНК в зв'язці U4-U6 і таким чином спрощувати формування зв'язку U2-U6.
Зв'язки між шпильками I і II в мяРНП U4 і U6 розриваються, і ділянка шпильки II U6, що звільнилася, згортається сама на собі з утворенням внутрішньомолекулярної шпильки. Після цього потреба в U4 відпадає. Ділянка шпильки I U6, що звільнилася, зв'язується з мяРНК U2 з утворенням U2-U6-спіралі I. Структура спіралі I, проте, взаємовиключна з 3'-половиною ділянки внутрішньої 5’-шпильки мяРНК U2.
Динаміка РНК-РНК взаємодій у сплайсосомі
Хоча точну роль багатьох окремих складових сплайсосоми ще належить визначити, їх основною функцією є організація складної серії мяРНК /пре-мРНК і мяРНК /мяРНК взаємодій.
Після завершення збірки сплайсосоми, U1 snRNP є основним в парі із 50-сплайсинг сайтом, U2 snRNP є основним в парі з точкою розгалуження, U5 snRNP вступає в контакт з 50 екзоном, U4 і U6 мяРНП; вони з'єднуються між собою за допомогою великої кількості зв'язків.
Протягом першого перегрупування, базове сполучення між U1 мяРНК і ділянкою сплайсингу 50 порушується і замінюється взаємодією спарювання основ між U6 мяРНК і 50 сайту сплайсингу. Одночасно з цим, U4 і U6 розмотуються (деконденсуються) і U6 мяРНК утворює багато основних взаємодій з U2 мяРНК.
Велика кількість досліждень доводить самочинність мяРНК-опосередкованого каталізу. Дійсно, фрагменти U2 мяРНК і U6 мяРНК здатні каталізувати реакцію, аналогічну першому ступеню сплайсингу в повній відсутності кофакторів відповідних білка[16].
Як бачимо, сплайсосома, як і рибосома, є в рибозимом.
Примітки
- Berget et al., 1977
- Chow et al., 1977
- Ezkurdia, I.; Juan, D.; Rodriguez, J. M.; Frankish, A.; Diekhans, M.; Harrow, J.; Vazquez, J.; Valencia, A. та ін. (2014). Multiple evidence strands suggest that there may be as few as 19 000 human protein-coding genes. Human Molecular Genetics 23 (22): 5866–5878. ISSN 0964-6906. doi:10.1093/hmg/ddu309.
- UniProtKB results: Homo sapiens Архівовано 18 серпня 2016 у Wayback Machine.(англ.)
- Jamison et al., 1992
- Seraphin and Rosbash, 1989
- Legrain et al., 1988
- Query et al., 1994
- Newby and Greenbaum, 2002
- Burge et al., 1999
- Staley and Guthrie, 1998
- Newman et al., 1995
- Chiara et al., 1997
- Moore et al., 1993
- Konforti et al., 1993
- Timothy W., Nilsen (2013). Encyclopedia of Biological Chemistry - Vol. 4. (English). с. 88–98.
Джерела
- Chapter 12, pp 311 7th ed, Vishal.
- Alberts, Bruce. Bray, Dennis. Hookin, Karen. Johnson, Alexander, Lewis, Julian, Raff, Martin, Roberts, Keith. Walter, Peter. essential cell biology Second edition, GS Garland Science, Taylor & Francis Group, NEW YORK AND LONDON.
Зовнішні посилання
- Nilsen T (2003). The spliceosome: the most complex macromolecular machine in the cell?. Bioessays 25 (12): 1147–9. PMID 14635248.
