Покритонасінні
Покритонасінні, або квіткові рослини (Angiospermae, або Magnoliophyta), зав'язко́ві, вкритонасі́нні[1] — таксон вищих рослин, які формують квітки. Ця група налічує 64 порядки, понад 400 родин, понад 13 000 родів і, ймовірно, не менше 250 000 видів сучасних рослин. За числом видів квіткові рослини значно перевершують решту груп вищих рослин, узятих разом. Вони складають одну з груп насінних рослин. Найважливіша особливість квіткових рослин — наявність спеціалізованого органу, квітки, що бере на себе функції розмноження і залучення агентів запилення. Квіткові рослини формують свої насінні зачатки в порожнині зав'язі, яка утворена зрощенням колись відкритого плодолистка. Стінки зав'язі після запліднення розростаються і видозмінюються, даючи структуру під назвою плід. У решті груп насінних рослин насіннєвий зачаток не прихований від запилення, а насіння не поміщене в дійсний плід, але іноді насіння можуть покривати м'ясисті структури, наприклад, у представників роду тис.
| Покритонасінні | |
|---|---|
 | |
| Квітка магнолії | |
| Біологічна класифікація | |
| Царство: | Рослини (Plantae) |
| Клада: | Судинні рослини (Tracheophyta) |
| Клада: | Насінні (Spermatophytes) |
| Клада: | Покритонасінні (Angiosperms) |
| Порядки | |
| |
Історія
Ботанічний термін «Angiospermae» (дослівно «покритонасінні») запропонував Пауль Герман (нім. Paul Hermann) в 1690 році, термін був складений з грецьких слів αγγειον (вмістище) і σπερμα (насіння). Так Герман називав один з найголовніших відділів рослинного царства, якій включав рослини, насіння яких оточене капсулами. Gymnospermae за Германом були квітковими рослинами, чий єдиний цілісний плід вважався насінням з відсутніми покривами. Сам термін і його антонім були підхоплені Карлом Ліннеєм, який використовував їх у схожому, але більш обмеженому сенсі — для назв рядів свого класу Двосильних (Didynamia). У своєму сучасному значенні ці терміни почали використовуватися після того, як Роберт Браун в 1827 році встановив існування істинно голих насіннєвих зачатків у Cycadeae і Coniferae, назвавши їх Gymnospermae. З того часу термін Angiospermae став використовуватися різними авторами, іноді з різними варіаціями, для позначення однієї з підгруп в межах дводольних рослин (розділення «явношлюбних» рослин на безсім'ядольні, одно(сім'я)дольні і дво(сім'я)дольні набуло поширення дещо раніше).
Проте після того, як Вільгельм Гофмейстер описав процеси, які відбуваються в зародковому мішку квіткових рослин (1851), і зіставив їх із заплідненням таємношлюбних, стало ясно, що Gymnospermae є групою абсолютно відмінною від Angiospermae. В результаті, поняття «покритонасінні» поступово стали розглядати як синонім поняття «квіткові», і, відповідно, дводольні (Magnoliopsida, або Dicotyledones) і однодольні (Liliopsida, або Monocotyledones) — як підгрупи у складі Angiospermae. У цьому значенні поняття «покритонасінні» (Angiospermae) використовується й донині.
Походження
Один з найважливіших напрямків розвитку рослинного царства — пристосування до мінливостей навколишнього середовища. Квіткові рослини є яскравим прикладом цієї лінії і домінують на земній поверхні в дану епоху. Від полюсів до екватора немає такої ділянки, де можливе рослинне життя, але не знайдено покритонасінних. Вони удосталь зустрічаються в долинах річок і чистих озерах, в меншій кількості — в солоних озерах і морях. Проте такі водні покритонасінні не є примітивними формами, а виникли шляхом пристосування наземного предка до водного середовища. До щонайширшої географічної різноманітності додається різноманітність форм і способів зростання. Банальна ряска, що покриває поверхню ставка, є крихітним зеленим пагоном з простим корінцем, вертикально зануреним у воду, і з дуже нечіткими листами і частинами стебла. Могутнє лісове дерево сторіччями розвивало свою складну систему стовбурів і гілок, покритих незліченними гілочками і листям, а під землею відповідну площу займає могутня, добре розвинена коренева система. Між цими двома крайнощами — нескінченні градації: водні і земні трави, повзучі, висхідні, кущі і дерева, набагато більша різноманітність, ніж серед решти груп насінних рослин, загалом відомих як голонасінні (Gymnospermae)[2].
Еволюція квіткових відбувалася швидко шляхом широкої адаптивної радіації. Перші залишки покритонасінних датують юрським періодом, тому вони мають вік приблизно 140 мільйонів років. Базуючись на сучасних даних, можна припустити, що предки покритонасінних і гнетових розділилися протягом тріасового періоду (220—202 мільйонів років тому)[3]. Залишки рослин з ознаками покритонасінних з'явилися протягом юрського і раннього крейдового періоду (135—65 мільйонів років тому), але це були досить-таки нечисленні і примітивні форми. Найдавнішою відомою покритонасінною рослиною є Montsechia vidalii, віком 130 млн років.[4] Сліди широкого розвитку і розповсюдження покритонасінних з'явилися в палеонтологічному літописі в середині крейдового періоду (близько 100 мільйонів років тому). А вже наприкінці цього періоду покритонасінні стали панівною формою рослинного життя, і в багатьох викопних рослинах пізнаються представники сучасних родин (наприклад, бук, дуб, клен і магнолія).
На відміну від ссавців, що мають яскраві приклади перехідних видів від плазунів такі як качкодзьоб і єхидна, квіткові рослини не мають жодного виду, про який можна сказати, що він займає проміжне положення між покритонасінними та іншими відділами вищих рослин.
Наявність насіння зближує їх з голонасінними. Деякі науковці вважали предками квіткових рослин ряд бенетити, оскільки вони єдині мали двостатеві шишки. Проте зараз цю гіпотезу спростували. Наразі вчені вважають, що квіткові рослини на початку розвитку були тісно пов'язані з насінними папоротями.
Ще Чарлз Дарвін припускав, що квіткові рослини спочатку розвивалися на обмеженій території, звідки вони завдяки географічним змінам змогли вирватися і швидко розповсюдитися по світу. Науковці А. Тахтаджян, Р. Торн, А. Сміт вважають, що цим місцем була Південно-Східна Азія. Там збереглися найпримітивніші родини: магнолієві, деґенерієві, вінтерові.
Перші квіткові рослини очевидно були чагарниками або невисокими деревами. Ксилема не мала судин. Листя вічнозелені з перистим жилкуванням. Квітки однодомні без пелюсток, що розвинулися потім. Запилювалися жуками. Плід — багатолистянка[5].
Одна з гілок квіткових рослин, а саме Частухоцвіті (близько 72 видів Posidoniaceae, Zosteraceae, Hydrocharitaceae і Cymodoceaceae), коло 100 млн років тому зуміли заново колонізувати морське дно, утворюючи специфічну екосистему — луки морських трав.[6]
Анатомія квіткових рослин
Квітка
- Основні статті: Квітка і Статеве розмноження квіткових рослин
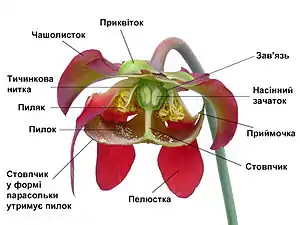
Характерною ознакою покритонасінних є квітка, яка демонструє чудову різноманітність форм і забезпечує надійні зовнішні ознаки для розрізнення видів покритонасінних. Функція квітки — забезпечення запліднення яйцеклітини і розвитку плоду, який містить насіння. Іноді, як у фіалки, квітка виникає окремо в пазусі звичайного листка. Проте, звичайно, частина рослини, що несе квітку, знаходиться на кінці паростка, видається над вегетативною частиною (частиною, яка несе листя) і формує детально розроблену систему гілок, відому як суцвіття.
Відтворювальні (репродуктивні) клітини покритонасінних можуть бути двох видів. Одні — мікроспори, або зерна пилку, є «чоловічими» клітинами і формуються в тичинках (мікроспорофілах). Інші — мегаспори, «жіночі» клітини, в яких розвивається яйцеклітина, містяться в насінному зачатку, котрий знаходиться в плодолистку (мегаспорофілі). Квітка може складатися тільки з цих частин, як у верби, де кожна квітка містить тільки декілька тичинок або два плодолистки. Однак зазвичай у квітці також є й стерильні структури, пристосовані як для захисту, так і для приваблення комах, необхідних для запилення. Зовнішня структура називається чашечкою і поділяється на чашолистки, які звичайно зелені та нагадують листя. Головна функція цих структур — захист квітки, особливо на стадії бруньки. Внутрішня структура називається віночком і складається з пелюсток, які загалом мають яскраві кольори і делікатнішу будову. Її функція полягає в привабленні птахів та комах — посередників, за допомогою яких відбувається запилення. Механізми цього приваблення звичайно включають також привабливий запах та нектар, який виділяється у квітці. Ці характеристики, які сформувалися у процесі еволюції для приваблення тварин-запилювачів, роблять квіти такими популярними серед людей. У деяких рослин, наприклад, магнолії, чашолистки і пелюстки не відрізняються одні від одних, у такому разі всі вони називаються чашолистками.
Тоді як більшість квіток повноцінні, або гермафродити, тобто містять як чоловічі, так і жіночі частини в одній структурі, квіткові рослини розвинули численні морфологічні і фізіологічні механізми, щоб зменшити можливість або запобігти самозаплідненню. Гетероморфні квіти мають короткі плодолистки і довгі тичинки або навпаки, щоб перешкодити тваринам-запилювачам занести пилок в маточку (сприйнятливу частину плодолистка). Гомоморфні квіти використовують біохімічний (фізіологічний) механізм, який називається само-несумісністю, щоб відрізнити свій пилок від чужого. У деякий видів чоловічі і жіночі частини морфологічно відокремлені, розвиваючись на різних квітках.
Запліднення і ембріогенез
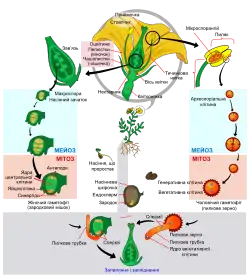
- Основні статті: Запліднення і Ембріогенез рослин
Подвійне запліднення — процес у квіткових рослин протягом відтворення, при якому один спермій зливається з яйцеклітиною, а другий — з диплоїдною центральною клітиною. Зерно пилку приклеюється до приймочки й вирощує пилкову трубку, яка проникає у насінний зачаток через крихітну пору, мікропіле. Одна з двох клітин сперми запліднює яйцеклітину, формуючи диплоїдну зиготу або ембріон, також відому як насінний зачаток. Друга клітини сперми зливається з двома гаплоїдними полярними ядрами в центрі ембріонального мішечка. В результаті формується триплоїдна клітина (3n). Ця клітина поділяється через мітоз і формує ендосперм — багату на поживні речовини тканину насіння. Якщо насіння розвивається без запліднення, цей процес називається апомікс.
Плід і насіння
Протягом того, як розвиток ембріона і ендосперми протікає в межах мішечка ембріона, його стіна збільшується і звичайно поглинає ядро (яке також збільшується), формуючи оболонку насіння. Стінка насінного зачатку також розвивається, формуючи плід, структуру, тісно пов'язану з розповсюдженням насіння. Часто запліднення впливає на інші частини квітки, які беруть участь в утворенні плоду, як квітколоже в яблуці, суницях та інших. Функція оболонки насіння подвійна — захисту ембріона і допомоги в розповсюдженні, вони також можуть безпосередньо допомагати проростанню. Якщо плід розкривається і тому насіння буде залишене незахищеним, оболонка забезпечує захист ембріона і, можливо, також розповсюдження. Якщо плід не розкривається, оболонка насіння майже не розвивається.
Таксономія

Квіткові рослини зараз розглядаються як невизначена таксономічна категорія рангом між відділом та класом. Оскільки ця таксономічна категорія більш високого рангу, ніж родина, є певна свобода у виборі назви. Стаття 16 Міжнародного кодексу ботанічної номенклатури (ICBN) дозволяє використовувати як і традиційні історичні назви, так і назву, утворену від роду. Офіційна уніномінальна назва цього таксона — Magnoliophyta, від назви роду Магнолія (Magnolia). Але традиційно укорінилися такі імена як Angiospermae і Anthophyta (квіткові рослини).
Завдяки постійному перегляду поглядів на спорідненість квіткових рослин, внутрішня систематика цієї групи піддавалася і піддається змінам. Дві широко використовувані, хоч і дещо застарілі, системи квіткових рослин — система Тахтаджяна і система Кронквіста. На сьогодні обидві ці класифікації не відображають філогенію таксона. Сучасна визнана класифікація створена міжнародною «Групою філогенії Покритонасінних» (англ. Angiosperm Phylogeny Group), яка опублікувала свій перший варіант таксономії 1998 (відомий як APG-I), другий 2003 (APG-II), третій 2009 (APG-III), четвертий 2016 року (APG-IV).
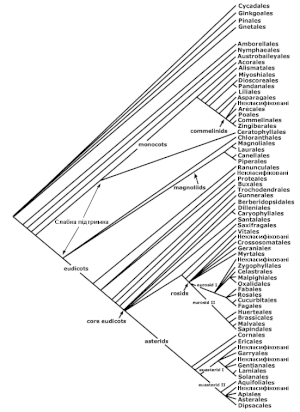
Традиційно, квіткові рослини були розбиті на дві групи, які в системі Кронквіста названі Magnoliopsida (у ранзі класу, сформованого від фамільного імені Magnoliacae) і Liliopsida (у ранзі класу, сформованого від фамільного імені Liliaceae). Інші описові імена, дозволені згідно зі статтею 16 ICBN, включають Dicotyledones або Dicotyledoneae, і Monocotyledones або Monocotyledoneae, які мають довгу історію використання. Для членів першої групи також може використовуватися термін «дводольна рослина», для другої — «однодольна», або «дікот» та «монокот» відповідно. Ці назви походять від факту, що дводольні звичайно мають у кожній насінині дві сім'ядолі (ембріональні листки), тоді як однодольні звичайно мають тільки одну. З діагностичної точки зору число сім'ядоль не є ні особливо зручною, ні надійною ознакою.
Недавні дослідження, проведені групою APG, показали, що однодольні — «хороша» (тобто монофілетична) група, цей таксон отримав назву «монокоти» (monocots), а дводольні — ні (вони створюють парафілетичну групу). Проте, в межах дводольних існує «хороша» група, яка включає більшість дводольних. Цей таксон називається «евдикоти» (eudicots) або «tricolpates» (від виду пилку, знайденого у представників цієї групи). Назва eudicots походить від слова «dicot» з префіксом «eu-» (від грецької 'eu'= «істина»), тобто означає «справжні дводольні», оскільки евдикоти мають ознаки, традиційні для дікотів, наприклад квіти з чотирьох або п'яти частин (чотирьох або п'яти пелюсток, чотирьох або п'яти чашолистків). Групу, яка залишається після відділення евдикотів з решти (колишніх) дікотів, іноді неофіційно називають «палеодікоти» (palaeodicots) (грецький префікс «palaeo-» означає «старий»). Оскільки представники цього залишку не створюють «хорошу» групу, цей термін існує тільки для зручності.
Квіткові рослини налічують приблизно 352 тисячі видів. Найбільше видів квіткових рослин містять родини айстрових (Asteraceae або Compositae), орхідних (Orchidaceae) і бобових (Fabaceae)[7]; найбільші роди — астрагал (Astragalus) і перець (Piper)[8]. Однак оцінка різноманітності груп рослин за кількістю видів не є однозначною. Якщо взяти зокрема родину зозулинцевих, то вони є сестринськими до решти холодкоцвітих і різниця в кількості видів між ними чотирикратна, тобто не така й значна. Своєю чергою холодкоцвіті є сестринськими до комелінідів, й різниця в кількості видів між ними зовсім незначна[9].
Економічна важливість
Сільське господарство майже повністю залежить від покритонасінних, або безпосередньо, або непрямо через харчування худоби. З родин квіткових рослин, злакові (Poaceae) — безумовно найголовніша, оскільки забезпечує більшу частину всіх вимог сільського господарства (рис, кукурудза, пшениця, ячмінь, жито, овес, просо, цукрова тростина, сорго), з бобовими (Fabaceae) на другому місці. Також високу важливість мають — пасльонові (Solanaceae): картопля, помідори, перець; гарбузові (Cucurbitaceae): кавуни, гарбузи і дині; капустяні (Brassicaceae): ріпак і капуста та окружкові (Apiaceae): петрушка. Багато плодів отримують від родин рутових (Rutaceae): помаранчі, цитрини, мандарини і розових (Rosaceae): яблука, груші, вишні, абрикоси, сливи тощо.
У деяких частинах світу єдиний вид набуває першорядного значення через різноманітність використань. Приклад — кокосовий горіх (Cocos nucifera) на тихоокеанських атолах. Інший приклад — маслина (Olea europaea) у Середземномор'ї.
Квіткові рослини також є постачальниками деревини, паперу, волокна (бавовник, льон і коноплі та ін.), ліків (наперстянка, камфора), використовуються як декоративні та мають інші застосування.
Див. також
Примітки
- Російсько-український академічний словник А. Кримський, С. Єфремов 1924—1933
- Як квіти завоювали світ. BBC Україна. 15 січня, 2018 рік.
- Проблеми походження покритонасінних
- Bernard Gomez, Véronique Daviero-Gomez, Clément Coiffard, Carles Martín-Closas, and David L. Dilcher. Montsechia, an ancient aquatic angiosperm.: — Proceedings of the National Academy of Sciences. August 17, 2015
- Происхождение цветковых растений // Жизнь растений. В 6-ти т. Т. 6. Цветковые растения / Под ред. А. Л. Тахтаджяна. — М.: Просвещение, 1982. — С. 103—107.
- Seagrass and Seagrass Beds | Smithsonian Ocean. ocean.si.edu (англ.). Процитовано 29 жовтня 2021.
- Armstrong W. P. Diversity Of Flowering Plants. Процитовано 29.12.2020.
- Plants of the World Online. Kew Science. Процитовано 29.12.2020.
- Stevens P. F. (2017). Angiosperm Phylogeny Website. Процитовано 29.12.2020.
Література
- Дендрофлора України. Дикорослі й культивовані дерева і кущі. Покритонасінні: Довідник. Ч. 1/ М. А. Кохно, Л. І. Пархоменко., А. У. Зарубенко та ін.; — Київ: Фітосоціоцентр, 2003. — 451 с.
- Дендрофлора України. Дикорослі і культивовані дерева і кущі. Покритонасінні: Довідник. Ч. 2 / М. А. Кохно, Н. М. Трофименко, Л. І. Пархоменко та ін. — К.: Фітосоціоцентр, 2005. — 716 с.
- (рос.) Красилов В. А. Происхождение и ранняя эволюция цветковых растений. — М. : Наука, 1994.
Посилання
- Покритонасінні рослини // Фармацевтична енциклопедія
- Angiosperm Phylogeny
- Angiosperm Phylogeny Group (2003). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141: 399—436. онлайн.
- Angiosperms — Tree of Life Web Project
- Angiosperms An annotated link directory
- Сікура Й. Й. Морфологічні особливості плодів та насіння квіткових рослин світової флори. — Ужгород: TIMPANI, 2014—376 с. (угорською та українською мовою) ISBN 978-966-8904-74-5
- Характеристика класів і окремих родин. Значення покритонасінних рослин у природі й у житті людини
