ДНК-зв'язувальні білки
ДНК-зв’язуючі білки [3][4][5] - це Білки, які утворені з ДНК-зв’язуючих доменів, а отже мають специфічну чи загальну спорідненість до одно- або двониткових ДНК. ДНК-зв’язуючі білки, які є сайт-специфічними, в основному взаємодіють із великою борозенкою B-ДНКа, тому що вона експонує більше функціональних груп, які ідентифікують пару основ. Однак відомі і до малої борозни ДНК-зв’язуючі ліганди, такі як Нетропсин, [6] Дістаміцин, Hoechst 33258, Пентамідин, ДАФІ(4.6-диамідо-2-фенол-індол) та інші.[7]

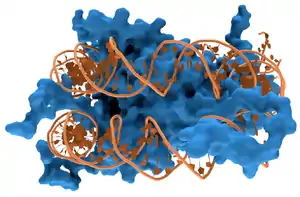
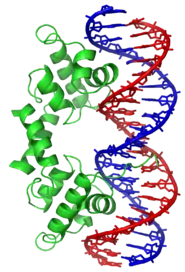
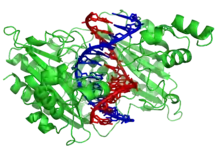
Приклади
ДНК-зв’язуючі Білки включають Фактори транскрипції які є генними модуляторами процесу транскрипції, різні Полімерази, Нуклеази які розрізають молекули ДНК, та Гістони які берутьь участь в компресії хромосом і транскрипції в клітинному ядрі. ДНК-зв’язуючі протеїни можуть містити такі домени, як цинковий палець, «спіраль-петля-спіраль», «лейцинова застібка» (одні з багатьох) , що сприяють зв’язуванні молекули до нуклеотиду.
Не специфічні ДНК-білкові взаємодії
Структурні білки, що зв’язують ДНК є добре вивченими прикладами неспецифічної ДНК-білкової взаємодії. В самій хромосомі ДНК зберігається в комплексі зі структурними білками. Ці білки впорядковують ДНК в компактну структуру під назвою Хроматин. В еукаріотів в цих структурах ДНК зв’язується з комплексом невеликих основних білківГістони, коли ж у прокаріотів залучається значна кількість протеїнів.[8][9] Гістони створюють диско-подібний комплекс під Нуклеосома, який містить два повні витки дво-ланцюгової ДНК, намотаної на її поверхні. Ці не-специфічні взаємодії формуються за рахунок основних груп в гістонах, створюючи іонні зв’язки з кислотними вуглеводно-фосфатними залишками хребта ДНК, і є, таким чином, значною мірою, незалежні від послідовності основ ДНК.[10] Хімічні модифікація цих амінокислотних залишків включає Метилювання, Фосфорилювання та Ацетилювання.[11] Ці хімічні модифікації змінюють міцність взаємодії між гістонами і ДНК, роблячи ДНК більш чи менш доступною для факторів транскрипції і змінюючи рівень транскрипції.[12] Інші не-специфічні ДНК-зв’язуючі протеїни в хроматині містять включають високо-мобільні ггрупи білків, які приєднюються, щоб натягнути чи деформувати ДНК.[13] Ці протеїни є важливими в згинанні масивів нуклеосом і впорядкування їх в більші структури, щоб утворити хромосоми. [14]
ДНК-зв’язуючі білки, що специфічно зв’язують до одно-ланцюгових молекул
Серед ДНК-зв’язуючих білків є окрема група молекул, які специфічно зв’язують одно-ниткові молекули ДНК. В людей, реплікативний протеїн А(RPA eng.) є найбільш вивчений член цієї родини білків. Він використовується в процесі де розділяється подвійна спіраль, включаючи ДНК реплікацію, рекомбінацію і ДНК репарацію. [15] Швидше за все дані білки стабілізують одно-ниткові ДНК і захищають їх від утворення шпильок чи деградації нуклеазами.
Зв’язування до специфічних ДНК послідовностей
На противагу, інші білки задіяні в зв’язуванні специфічних послідовностей. Найбільш інтенисивно вивчаються різні Фактори транскрипції, які є білками, що регулюють. Кожен транскрипційний фактор зв’язкується до однієї специфічної множини ДНК послідовностей і активує чи інгібує транскрипцію генів, які містять дані послідовності біля своїх промоуторів. Фактори транскрипції роблять це двома шляхами. По-перше, вони можуть приєднюватися до РНА-полімерази відповідальної за транскрипцію або прямо, або через інший медіаторний білок, це змінює локацію полімерази, і вона наближається до промотора, і дозволяє їй розпочати процес тракнскрипції. [16] Або ж, фактор транскрипції може зв’язувати Ферменти, що модифікують гістони біля промоторної ділянки; і це змінить рівень доступності ДНК матриці для полімерази.[17]
Так як ці ДНК-цілі можуть опинитися будь-де в людському геномі, зміна в активностьі одного типу фактора транксрипці може вплинути на тисячі генів.[18] Як наслідок, ці білки є часто мішенею для Трансдукції сигналу, процесу що контролює відповідь на зміну зовнішнього середовища чи диференціювання клітині їх розвиток. Специфічність цих факторів транскрипції взаємодії з ДНК походить від того, білки створють декілька контактів з гранями основ ДНК, що й дозволяє ім «прочитати»ДНК послідовність. Більшість цих взаємодій відбувають у великій борозенці, де основи є більше доступні. [19] Математичний опис білок-ДНК зв’язування враховує спеціфічність послідовності, конкурентність та кооперацію в процесі зв’язування білків різних типів, і зазвичай здійснюється з допомогою біофізичного.[20]
Посилання
- Created from PDB 1LMB
- Created from PDB 1RVA
- Travers, A. A. (1993). DNA-protein interactions. London: Springer. ISBN 978-0-412-25990-6.
- Pabo CO, Sauer RT (1984). Protein-DNA recognition. Annu. Rev. Biochem. 53 (1): 293–321. PMID 6236744. doi:10.1146/annurev.bi.53.070184.001453.
- Dickerson R.E. (1983). The DNA helix and how it is read. Sci Am 249 (6): 94–111. doi:10.1038/scientificamerican1283-94.
- Zimmer C, Wähnert U (1986). Nonintercalating DNA-binding ligands: specificity of the interaction and their use as tools in biophysical, biochemical and biological investigations of the genetic material. Prog. Biophys. Mol. Biol. 47 (1): 31–112. PMID 2422697. doi:10.1016/0079-6107(86)90005-2.
- Dervan PB (April 1986). Design of sequence-specific DNA-binding molecules. Science 232 (4749): 464–71. PMID 2421408. doi:10.1126/science.2421408.
- Sandman K, Pereira S, Reeve J (1998). Diversity of prokaryotic chromosomal proteins and the origin of the nucleosome. Cell Mol Life Sci 54 (12): 1350–64. PMID 9893710. doi:10.1007/s000180050259.
- Dame RT (2005). The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin. Mol. Microbiol. 56 (4): 858–70. PMID 15853876. doi:10.1111/j.1365-2958.2005.04598.x.
- Luger K, Mäder A, Richmond R, Sargent D, Richmond T (1997). Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389 (6648): 251–60. PMID 9305837. doi:10.1038/38444.
- Jenuwein T, Allis C (2001). Translating the histone code. Science 293 (5532): 1074–80. PMID 11498575. doi:10.1126/science.1063127.
- Ito T (2003). Nucleosome assembly and remodelling. Curr Top Microbiol Immunol 274: 1–22. PMID 12596902. doi:10.1007/978-3-642-55747-7_1.
- Thomas J (2001). HMG1 and 2: architectural DNA-binding proteins. Biochem Soc Trans 29 (Pt 4): 395–401. PMID 11497996. doi:10.1042/BST0290395.
- Grosschedl R, Giese K, Pagel J (1994). HMG domain proteins: architectural elements in the assembly of nucleoprotein structures. Trends Genet 10 (3): 94–100. PMID 8178371. doi:10.1016/0168-9525(94)90232-1.
- Iftode C, Daniely Y, Borowiec J (1999). Replication protein A (RPA): the eukaryotic SSB. Crit Rev Biochem Mol Biol 34 (3): 141–80. PMID 10473346. doi:10.1080/10409239991209255.
- Myers L, Kornberg R (2000). Mediator of transcriptional regulation. Annu Rev Biochem 69 (1): 729–49. PMID 10966474. doi:10.1146/annurev.biochem.69.1.729.
- Spiegelman B, Heinrich R (2004). Biological control throughs regulated transcriptional coactivators. Cell 119 (2): 157–67. PMID 15479634. doi:10.1016/j.cell.2004.09.037.
- Li Z, Van Calcar S, Qu C, Cavenee W, Zhang M, Ren B (2003). A global transcriptional regulatory role for c-Myc in Burkitt's lymphoma cells. Proc Natl Acad Sci USA 100 (14): 8164–9. PMC 166200. PMID 12808131. doi:10.1073/pnas.1332764100.
- Pabo C, Sauer R (1984). Protein-DNA recognition. Annu Rev Biochem 53 (1): 293–321. PMID 6236744. doi:10.1146/annurev.bi.53.070184.001453.
- Teif V.B., Rippe K. (2010). Statistical-mechanical lattice models for protein-DNA binding in chromatin.. Journal of Physics: Condensed Matter. arXiv:1004.5514.
Зовнішні посилання
- Abalone tool for modeling DNA-ligand interactions.
- DBD database of predicted transcription factors Uses a curated set of DNA-binding domains to predict transcription factors in all completely sequenced genomes
- MeSH DNA-Binding+Proteins