Нуклеосома
Нуклеосома — структурна частина хроматину, утворена ділянкою нитки ДНК намотаною на серцевину із основних білків-гістонів, має діаметр ~11 нм[1]. Нуклеосоми є першим рівнем упакування ДНК еукаріот (необхідне для поміщення ДНК загальною довжиною близько метра у ядро діаметром 5—10 мкм), а також архей, і забезпечує її компактизацію приблизно у сім разів і захищає її від ушкоджень[2]. Серцевина нуклеосоми складається із восьми білків: по дві копії гістонів H2A, H2B, H3 і H4, N-кінець (хвіст гістону) кожної білкової молекули виступає назовні нуклеосоми і може бути місцем різних ковалентних модифікацій. Довжина ділянки ДНК, що входить до складу нуклеосоми, становить 146 п.н., вона намотана на білкову серцевину у вигляді лівозакрученої суперспіралі 1,7 рази. Нуклеосоми розміщуються вздовж ДНК досить регулярно, на найчастіше відстані від 8 до 80 (довжина лінкерної ДНК, зв'язаної із гістоном H1)[3]. Таким чином нуклеосоми повторюються приблизно кожних 200 п.н., одна диплоїдна клітина людини містить приблизно 30 млн нуклеосом[1]. Послідовність нуклеосом і лінкерних ділянок формує 10 нм філамент, що має вигляд «намистин на нитці» і переважно підлягає подальшій компактизації.
.png.webp)
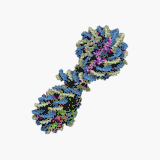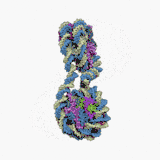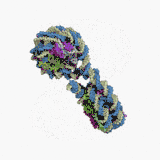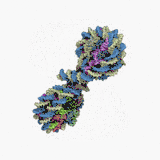
Технічно, термін «нуклеосома» позначає ќорову частинку (146 п.н. ДНК і білкову серцевину) і одну найближчу лінкерну послідовність, але переважно вживається як синонім корової частинки[1].
Впродовж клітинного циклу нуклеосоми тимчасово від'єднуються від ДНК тільки під час реплікації, також вони можуть бути усунуті на ділянках, що транскрибуються у певний момент. Додатково в сперматозоїдах під час їхнього дозрівання більшість нуклеосом заміщується на нуклеопротаміни, де основою є білки протаміни замість гістонів. Це додатково ущільнює ДНК, робить об'єм ядра сперматозоїда меншим та захищає ДНК від пошкоджень.[4]
Історія дослідження
Перші дані про особливості компактизації ДНК в ядрі еукаріот були отримані Морісом Вілкінсом у 1960-их роках. Він проводив рентгеноструктурний аналіз хроматину і з'ясував, що в ньому наявні певні повторювані елементи, які не спостерігаються ні в чистих препаратах ДНК, ані гістонів. Із цього Вілкінс зробив висновок, що гістони забезпечують впорядковану структурну організацію ДНК.
Перші дані електронної мікроскопії
1974 року Ада і Дональд Олінси опублікували електронні мікрографії волокон хроматину, що були виділені і підготовлені до мікроскопування м'якшими методами ніж, ті що використовувались раніше. Уникнувши застосування жорстких розчинників, вони змогли побачити структуру, що нагадувала «намистинки на нитці» — маленькі щільні тільця сполучені тонкими філаментами. Припускалось, що тільця, утворені білками, ймовірно гістонами, а філаменти — ниткою ДНК[2]. Олінси називали відркиті ними «намистини» «ν (ню) частинками», оскільки вони були новими і частиною нуклеогістну (комплексу ДНК і гістонами). Сам термін нуклеосома з'явився у 1975 року (P. Oudet, M. Gross-Bellard, P. Chambon)[5].
Незалежно від Олінсів того ж, 1974 року, структуру «намистини на нитці» вдалось візуалізувати за допомогою електронного мікроскопа також і Вудкоку, проте його стаття не була опублікована у Nature, рецензент відгукнувся про неї таким чином:
Еукаріотична хромосома, що складається із самозбірних 70 Å одиниць, які, мабуть, можливо кристалізувати, вимагала б переписування наших підручників із цитології і генетики! Я ніколи не бачив такої наївної статті, яка би претендувала на таку фундаментальну значимість. Однозначно, це не слід ніде публікувати! Оригінальний текст (англ.)A eukaryotic chromosome made out of self-assembling 70 Å units, which could perhaps be made to crystallize, would necessitate rewriting our textbooks on cytology and genetics! I have never read such a naive paper purporting to be of such fundamental significance. Definitely it should not be published anywhere!
Мікрографії Вудкока були опубліковані тільки 1976 року[5].
Фрагментація ДНК хроматину під дією нуклеаз
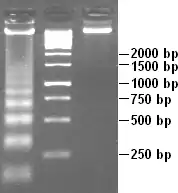
Приблизно в той же час були отримані інші результати, що вказували на існування у хроматині повторюваної структури, і підтверджували, що дані Олінсів були не просто артефактами приготування мікроперпаратів. Дін Г'юїш (англ. Dean Hewish) і Лі Бергойн (англ. Leigh Burgoyne) відкрили у ядрі гепатоцитів пацюків нуклеазу, здатну розщеплювати ДНК у нитках хроматину. В одному із експериментів після обробки хроматину цим ферментом, вони виділили частково деградовану ДНК і розділили її методом гель-електрофорезу. З'ясувалось, що найкоротші фрагменти ДНК мали довжину 200 , а довші були кратні 200 — 400, 600, 800 і так далі. Оскільки розщеплення цією ж нуклеазою ДНК чистої від білків не давало такої картини, Г'юїш і Бергойн зробили висновки, що білки хроматину утворюють структури вздовж ДНК, які повторюються кожні 200 п.н., і між цими структурами ДНК є чутливою до дії нуклеаз[2].
Щоб підтвердити взаємозв'язок між продуктами часткової деградації ДНК довжиною 200 п.н. і сферичними частинками, виявленими за допомогою електронної мікроскопії, використовувалась комбінація обидвох методів. Нитки хроматину спочатку обробляли нуклеазою мікрококів, отримані фрагменти розділяли за масою шляхом диференційного центрифугування. Кожну фракцію мікроскопували, а також виділяли із неї ДНК і визначали її розмір за допомогою гель-електрофорезу. З'ясувалось, що найлегша фракція містила поодинокі сферичні частинки і фрагменти ДНК довжиною ~200 п.н., наступна — по дві з'єднані частинки і ДНК довжиною ~400 п.н. і так далі. Таким чином стало відомо, що базовою структурною одиницею хроматину є нитка ДНК довжиною 200 п.н. намотана на серцевину із білків — нуклеосома[2].
Дослідження молекулярної структури нуклеосом
Подальші деталі молекулярної організації нуклеосом були з'ясовані завдяки працям групи Роджера Корнберга. Він показав, що структуру «намистини на нитці» можна отримати проінкубувавши очищену ДНК із димерами гістонів H2A-H2B і H3-H4. Щоб визначити скільки саме таких димерів входить до складу однієї нуклеосоми, Корнебрг разом із Джин Томас обробляли хроматин таким чином, що між усіма білковими молекулами, розташованими поруч, утворювались поперечні зшивки. Після цього комплекси зі зшивками виділяли і визначали молекулярну масу за допомогою гель-електрофорезу. Виявилось, що розміри отриманих комплексів відповідали восьми білкам-гістонам. Таким чином було встановлено, що нуклеосома складається зі двох димерів H2A-H2B і двох димерів H3-H4, тобто є октамером. Пізніше було показно, що гістон H1 асоційнований із лінкерними послідовностями і необхідний для вищих рівнів компактизації хроматину[2].
Відкриття нуклеосоми суттєво позначилось на уявленні про функціонування всього апарату реалізації і передачі спадкової інформації. ДНК вже не вважалась вкритою білками-гістонами, а навпаки — намотаною зовні на білокову серцевину. Таким чином з'ясувалось, що структура хроматину є більш відкритою до взаємодії із різноманітними факторами. Над кристалічною структурою нуклеосоми працювали відразу дві конкуруючі групи: Т. Річмонда і Г. Буніка. Зрештою вона була опублікована Річмондом і колегами у 1984 року із роздільністю 7,0 Ǻ, детальнішу структуру (2,8 Ǻ) розшифрували Каролін Люгер et al 1997 року[5].
Наступною важливою віхою у дослідженні нуклеосм було відкриття їхньої ролі в епігенетичній регуляції. Вперше гіпотеза про те, що епігенетична інформація може бути записана у формі модифікацій гістонових хвостів була запропонована 1993 року Тернером. Згодом з'явилась концепція «гістонового коду» (T. Jenuwein, C. Allis, 2001), яка дискутується і досі[5].
Структура нуклеосоми
Гістонова серцевина нуклеосоми має форму диска, на який намотана ДНК довжиною 146 н.м. у формі лівозакрученої спіралі. ДНК і білками у її складі утримуються великою кількістю нековалентних зв'язків, зокрема між фосфатними (негативно зарядженими) групами ДНК і позитивно зарядженими залишками амінокислот аргініну і лізину білків-гістонів, а також 142 водневими зв'язками і гідрофобними взаємодіями. N-кінець, або «хвіст» кожного із білків-гістонів виступає назовні корової частинки і може бути місцем ковалентної модифікації або/і взаємодії із іншими ядерними факторами[1][6].
Властивості ДНК у складі нуклеосоми

У складі нуклеосом ДНК характеризуєтьсня негативною суперскрученістю, через те що, щільне обертання цієї молекули навколо білкової серцевини потребує прибирання приблизно одного витка всередині самої спіралі ДНК. Наявність негативних суперспіралей у складі нуклеосом призводить до виникнення позитивних суперспіралей у лінкерних ділянках, останні можуть бути розслаблені еукаріотичними топоізомеразами[3].
Попри те, що більшість іонних і водневих зв'язків із білками утворює цукрово-фосфатний остов ДНК, внаслідок чого нуклеосома може формуватись на ДНК із будь-якою послідовністю нуклеотидів, остання все ж впливає на силу взаємодії. Вибірковість зв'язування гістонів із певними ділянками не до кінця зрозуміла, проте з'ясовано, що у місцях найщільнішого контакту між ДНК і білками переважно розташовані А=Т пари, через що стає можливим стискання малого жолоба ДНК, необхідне для цієї взаємодії. Дві або три пари А=Т підряд роблять зв'язування ще легшим. Через це in vivo близько половини нуклеосом розміщені у ділянках, де динуклеотиди АА, АТ або ТТ розкидані на відстані ~10 п.н.[3]. Проте селективність нуклеосом щодо послідовності нуклеотидів у ДНК є достатньо слабкою для того, щоб інші фактори домінували у виборі місця їх локалізації[1].
Формування нуклеосом
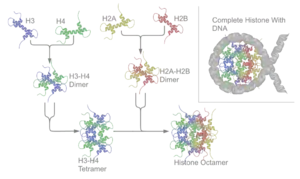
Нуклеосоми формуються відразу після реплікації ДНК, або інших процесів, що вимагають їх тимчасового усунення. Білкова серцевина нуклеосоми збирається у кілька етапів: спершу формуються димери H2A-H2B і H3-H4, після цього два димери H3-H4 об'єднуються у тетрамер, до якого згодом приєднуються два димери H2A-H2B[1]. Утворені білкові серцевини після реплікації включаються до хроматину за участі білкового комплексу RCAF (англ. replication-coupling assembly factor), що містить ацетильовані гістони H3 і H4, тримерний білок CHAF1A (англ. chromatin assembly factor 1) і білок ASF1A (англ. anti-silencing factor 1). Хоч механізм дії RCAF повністю не з'ясований, вважається, що він безпосередньо взаємодіє із апаратом реплікації. Якщо нуклеосоми повинні бути утворені після репарації або інших процесів, це забезпечується іншими білковими комплексами. Існують також фактори обміну гістонів, що можуть заміщувати гістони у нуклеосомі на варіантні форми[3].
Динаміка нуклеосом
Незважаючи на те, що ДНК і корові гістони утворюють міцно зв'язані нековалентними взаємодіями, нуклеосоми є досить динамічними структурами. Так кінетичні експерименти показали, що в ізольованій нуклеосомі ДНК розгортається із кожного кінця приблизно 4 рази на хвилину і залишається відкритою на 10—50 мс. У цей час вона може взаємодіяти із негістоновими білками, що є необхідним зокрема для транскрипції[1]. Також нуклеосоми обов'язково відділяються від ДНК під час таких процесів як реплікація та транскрипція, рекомбінація та репарація. Після завершення цих процесів нуклеосоми повертаються, але їхній склад може бути іншим. Так при реплікації ДНК з однієї материнської ДНК утворюється дві дочірніх ДНК, тому нуклеосом також треба вдівчі більше, що потребує нових гістонових білків. Проте після реплікації гістони "дочірніх" нуклеосом отримають більшість модифікацій гістонів "материнських" нуклеосом в тих же локусах, завдячуючи ще не до кінця відомим механізмам на 2017 рік[7]
У клітинах також наявні десятки різних компелксів ремоделювання хроматину, які можуть забезпечувати ковзання нуклеосом вздовж ДНК, «розбирати» їх або замінювати окремі білки-гістони. Такі комплекси складаються із 10 і більше субодиниць, є еволюційно спорідненими із ДНК-геліказами і працюють у АТФ-залежному режимі, тобто використовують для роботи енергію гідролізу цієї сполуки. Вони взаємодіють одночасно із коровими білками і ДНК, намотаною на них, і послаблюють зв'язки між цими компонентами нуклеосоми. Таким чином комплекси ремоделювання хроматину можуть «посувати» нуклеосоми, в результаті переводячи хроматин у більш відкритий, або навпаки — щільніше упакований стан. Окрім того, завдяки взаємодії із негативно зарядженими білками гістоновими шаперонами, комплекси ремоделювання хроматину можуть здійснювати обмін гістонів, наприклад, димерів H2A-H2B, або повністю усувати білкову серцевину. Діяльність кожного із цих білкових комплексів, що мають різне призначення, ретельно регулюється клітиною[1].
На розташування нуклеосом на нитці ДНК впливає велика кількість факторів, найважливішим серед яких є наявність інших білків приєднаних до молекули нуклеїнової кислоти. Деякі із них сприяють формуванню нуклеосом поблизу, інші навпаки — створюють для цього перешкоди[1].
Вільна від нуклеосом ділянка
В промоторній ділянці генів, також в активних енхансерах, існує ділянка ДНК, яка не зв'язана з нуклеосомами (англ. nucleosome-free region, NFR, чи англ. nucleosome-depleted region, NDR). Ця ділянка є важливою для зв'язування ДНК з транскрипційними факторами, приєднання факторів ініціації транскрипції та подальшої транскрипції гена. Вона обмежнована двома нуклеосомами: +1-нуклеосома, що знаходиться на досить сталій відстані від сайту старту транскрипції, та -1-нуклеосома, що розміщена в протилежному напрямку. Наступні декілька нуклеосом, як в плюс так і в мінус сторону, зазвичай також знаходяться на досить сталих позиціях, проте далі вони вже розмиваються і позиціювання нуклеосом всередині гену може бути різним.[7] Типова довжина вільної від нуклеосом ділянки — 140 нуклеотидів, 200 у дріжджів S. cerevisiae[8] Також фланкуючі цю ділянку нуклеосоми мають нетиповий склад гістонів, часто включають H2A.Z варіант гістону замість H2A, що трохи дестабілізує нуклеосому, полегшуючи її відділенню від ДНК.[7]
Роль нуклеосом в епігенетичній регуляції
- Див. також: Епігенетика
Гістони у складі нуклеосом можуть підлягати різним ковалентним модифікаціям (зокрема таким як метилювання, ацетилювання, фосфорилювання, АДФ-рибозилювання тощо) або/і заміщуватись на варіантні форми. Всі ці зміни впливають на властивості хроматину і можуть бути використані для регуляції експресії генів, позначення певних ділянок хромосом зі специфічними функціями (наприклад центромер), бути необхідними для таких процесів як репарація і генетична рекомбінація[3]. Модифікації гістонів можуть передаватись наступним поколінням клітин під час мітозу.
Варіантні гістони кодуються іншими генами, ніж основні типи, і на відміну від останніх, експресуються впродовж всього клітинного циклу і включаються до складу хроматину не залежно від процесу реплікації за участі гістонових шаперонів і компелксів ремоделювання хроматину[6]. Більшість варіантів були виявлені для гістонів H2A і H3. Наприклад, у ділянках еухроматину, які менш щільно упаковані і транскрипційно активні, в нуклеосомах використовуються варіанти H3.3 і H2A.Z. Останній стабілізує октамер гістонів у білковій серцевині нуклеосоми, проте одночасно пригнічує взаємодію між нуклеосомами, що є необхідною умовою для забезпечення вищих рівнів компактизації хроматину. Це призводить до більш відкритого стану хроматину[3].
Інший варіант гістону H2A — H2A.X пов'язаний із репапрацією і рекомбінацією ДНК. Нестача цього білка у мишей має наслідком генетичну нестабільність і чоловічу безплідність. Невеликі кількості нуклеосом, що містять H2AX розкидані по всьому геному, якщо поблизу такої нуклеосоми стається двонитковий розрив ДНК, H2AX фосфорилюється по залишку Ser139. Остання подія необхідна для збирання апарату репарації у цьому місці[3].
CENPA — ще один варіант гістону H3 пов'язаний із повторами ДНК у ділянках центромер. Миші без гену цього гістону нежиттєздатні[3].
Еволюція нуклеосом
У 1990-их роках було з'ясовано, що нуклеосоми наявні не тільки в еукаріот, а й в архей, проте в останніх вони містять не вісім, а чотири білки гістони (гомологи гістонів H3 і H4) і захищають приблизно 60 п.н. ДНК. Цілогеномний аналіз галофільної архебактрії Haloferax volcanii показав, що хроматин у цього організму організований надзвичайно схоже до еукаріотичного. Зокрема, було показано, що регуляторні послідовності на початку і кінці генів значно рідше зайняті нуклеосомами, ніж інші ділянки. Також з'ясувалось, що кількість нуклеосом, що припадає на одну й ту ж довжину ДНК, вдвічі більша в архей, ніж в еукаріот, що можна пояснити їхніми приблизно вдвічі меншими розмірами. Очевидно, що нуклеосоми виникли в процесі еволюції до розділення доменів архей та еукаріот. Проте, зважаючи на невеликі на те, що геном архебактрій представлений відносно невеликими кільцевими ДНК, які до того ж не обмежені ядерною мембраною, припускається, що первинною функцією нуклеосом була не компактизація, а участь у регуляції експресії генів[9].
Джерела
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. с. 211—217. ISBN 978-0-8153-4105-5.
- Hardin J, Bertoni G, Kleinsmith LJ (2011). Becker’s world of the cell (вид. 8th). Benjamin Cummings. с. 530—535. ISBN 0-321-71602-7.
- Nelson D.L., Cox M.M. (2008). Lehninger Principles of Biochemistry (вид. 5th). W. H. Freeman. с. 963—968. ISBN 978-0-7167-7108-1.
- Blanco, Mélina; Cocquet, Julie (2019). Genetic Factors Affecting Sperm Chromatin Structure. Advances in Experimental Medicine and Biology 1166. с. 1–28. ISSN 0065-2598. PMID 31301043. doi:10.1007/978-3-030-21664-1_1. Процитовано 27 лютого 2020.
- Olins D.E., Olins A.L. Chromatin history: our view from the bridge // Nature Reviews Molecular Cell Biology. — 2003. — Т. 4, вип. 10 (October). — С. 809—815. — DOI:. — PMID:. Процитовано 13.07.2013.
- Luger K, Dechassa ML, Tremethick DJ. New insights into nucleosome and chromatin structure: an ordered state or a disordered affair? // Nature Reviews Molecular Cell Biology. — 2012. — Т. 7, вип. 13 (July). — С. 436—47. — DOI:. — PMID:.
- Lai, William K. M.; Pugh, B. Franklin. Understanding nucleosome dynamics and their links to gene expression and DNA replication. Nature Reviews Molecular Cell Biology. doi:10.1038/nrm.2017.47.
- Yuan, Guo-Cheng; Liu, Yuen-Jong; Dion, Michael F.; Slack, Michael D.; Wu, Lani F.; Altschuler, Steven J.; Rando, Oliver J. (22 липня 2005). Genome-scale identification of nucleosome positions in S. cerevisiae. Science (New York, N.Y.) 309 (5734). с. 626–630. ISSN 1095-9203. PMID 15961632. doi:10.1126/science.1112178.
- Ammar R, Torti D, Tsui K, Gebbia M, Durbic T, Bader GD, Giaever G, Nislow C. Chromatin is an ancient innovation conserved between Archaea and Eukarya // eLife. — 2012. — December. — DOI:. — PMID:.
Посилання
- Goodsell, David (July 2000). Nucleosome. RCSB PDB. Molecule of the Month (англ.). Архів оригіналу за 6 липня 2013. Процитовано 5 липня 2013.
- Сиволоб А.В., Афанасьєва К.С. Молекулярна організація хромосом. с. 49—93.
- Epigenetics Overview. YouTube (англ.). CellSignal. 2011. Процитовано 16 липня 2013.
- X Inactivation and Epigenetics. YouTube (англ.). WEHImovies. 2012. Процитовано 16 липня 2013.
