Ацетилювання
Ацетилювання (або етаноїлювання відповідно до номенклатури IUPAC) — це реакція введення ацетильної функціональної групи до хімічної молекули. Деацетилювання — це видалення ацетильної групи.
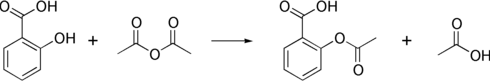
Ацетилювання стосується процесу введення ацетильної групи (в результаті чого утворюється ацетокси група) до молекули, а саме заміщення ацетильною групою протону водню. Реакцію з заміною протону водню в гідроксильній групі на ацетильну групу (СН3СО) утворює складний ефір, який називають ацетатом. Зазвичай використовують оцтовий ангідрид, як ацетилюючий агент, який реагує з вільними гідроксильними групами. Таким чином синтезують аспірин, героїн та THC-O-ацетат.
Ацетилювання білків
Ацетилювання — важлива модифікація білків у клітинній біології. Дослідження з протеоміки ідентифікували тисячі ацетильованих білків у ссавців.[1][2][3] Ацетилювання відбувається як ко-трансляційна та посттрансляційна модифікація білків шляхом ковалентного приєднання до них ацетильних груп, що відбивається на їх функціональній активності, наприклад, гістонів, p53 та тубулінів. Локальне ацетилювання N-кінцевих учасників гістонів на посттрансляційному рівні пов'язане із активацією хроматину. Специфічна ацетилаза (фермент, який каталізує процес ацетилювання) здійснює ацетилювання гістонів, беручи участь таким чином в регуляції транскрипції генів. Ацетилювання былка p53 по COOH-кінцю стимулює його здатність зв'язуватися із ДНК. Такі модифікації масово представлені серед хроматинових білків та ферментів клітинного метаболізму, що свідчить про те, що ацетилювання має значний вплив на експресію генів та метаболізм. У бактерій 90 % білків, які беруть участь у центральному обміні Salmonella enterica, ацетильовані.[4][5]
N-термінальне ацетилювання

N-кінцеве ацетилювання є однією з найпоширеніших ко-трансляційних ковалентних модифікацій амінокислот еукаріотів, яка відіграє важливу роль у регуляції функціонування різних білків. N-кінцеве ацетилювання визначає рівень синтезу, стабільність та локалізацію білків. Близько 85 % усіх білків людини і 68 % білків дріжджів ацетильовані на їх Nα-кінцях.[6] Кілька білків прокаріотів і археїв також модифікуються N-кінцевим ацетилюванням.
N-кінцеве ацетилювання каталізується набором ферментних комплексів, N-кінцевих ацетилтрансфераз (NATи). NATи переносять ацетильну групу з ацетил-коензиму А (Ac-CoA) до α-аміногрупи першого амінокислотного залишку білка. Різні NATи відповідають за ацетилювання N-кінцевого білка - амінокислотного ланцюга, який синтезується. Ацетилювання досі вважалося незворотним.[7]
N-кінцеві ацетилтрансферази
На сьогоднішній день у людини виявлено шість різних типів NAT — NatA, NatB, NatC, NatD, NatE та NatF. Кожен з цих різних ферментних комплексів є специфічним для різних амінокислот або амінокислотних послідовностей, які показані в наступній таблиці.
Таблиця 1. Склад та субстратна специфіка NAT.
| NAT | Субодиниці (каталітичні субодиниці виділені жирним шрифтом) |
Субстрати |
|---|---|---|
| NatA | Naa10 (Ard1) Naa15 (Nat1) | Ser-, Ala-, Gly-, Thr-, Val-, Cys- N-закінчення |
| NatB | Naa20 (Nat3) Naa25 (Mdm20) | Met-Glu-, Met-Asp-, Met-Asn-, Met-Gln- N-закінчення |
| NatC | Naa30 (Mak3) Naa35 (Mak10) Naa38 (Mak31) | Met-Leu-, Met-Ile-, Met-Trp-, Met-Phe- N-закінчення |
| NatD | Naa40 (Nat4) | Ser-Gly-Gly-, Ser-Gly-Arg- N-закінчення |
| NatE | Naa50 (Nat5) Naa10 (Ard1) Naa15 (Nat1) | Met-Leu-, Met-Ala-, Met-Lys-, Met-Met- N-закінчення |
| NatF | Naa60 | Met-Lys-, Met-Leu-, Met-Ile-, Met-Trp-, Met-Phe- N-закінчення |
| NatH | Naa80 | Актин- N-закінчення |
NatA
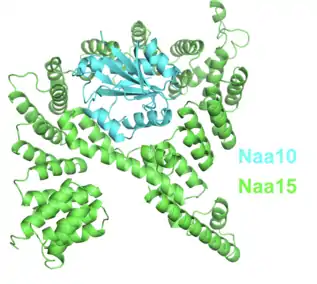
NatA складається з двох субодиниць: каталітичної субодиниці Naa10 та допоміжної субодиниці Naa15. Субодиниці NatA складніші у вищих еукаріотів, у порівнянні з нижчими еукаріотами. Naa10/Naa15 є найпоширенішим NatA і кодуються генами NAA10 та NAA15, відповідно. На додаток до генів NAA10 та NAA15, у ссавців є специфічні гени NAA11 і NAA16, яким відповідають функціональні білкові продукти, що утворюють різні активні комплекси NatA. Ці чотири каталітичні білки утворюють чотири можливі каталітично-допоміжні димери hNatA.[9]
NatA ацетилати Ser-, Ala-, Gly-, Thr-, Val- та Cys- N-кінцеві після того, як ініційований метіонін видаляється метіонинамінопептидазами. Ці амінокислоти частіше експресуються в N-кінцевих білках еукаріотів, тому NatA є основним NAT, що відповідає всій кількості його потенційних субстратів.[10]
Кілька різних партнерів по взаємодії беруть участь у N-кінцевому ацетилюванні NatA. Хантінтин-взаємодіючий білок К (HYPK) взаємодіє з hNatA на рибосомі для впливу на N-кінцеве ацетилювання підмножини субстратів NatA. Субодиниці hNaa10 та hNaa15 посилюють тенденцію до агрегації Хантінтіна, якщо HYPK виснажується. Було також встановлено, що індукований гіпоксією фактор (HIF)-1α взаємодіє з hNaa10, щоб інгібувати активацію опосередкованої hNaa10 активності транскрипції β-катеніну.[11]
NatB
Комплекси NatB складаються з каталітичної субодиниці Naa20p та допоміжної субодиниці Naa25p, які є як у дріжджів, так і у людини. У дріжджів всі субодиниці NatB пов'язані з рибосомами; а у людини субодиниці NatB виявляються як пов'язаними із рибосомою, так і у нерибосомальній формі. NatB ацетилює N-кінцевий метіонін субстратів, починаючи з терміналів Met- Glu-, Met-Asp-, Met-Asn- або Met-Gln- N-кінців.
NatC
Комплекс NatC складається з однієї каталітичної субодиниці Naa30p та двох допоміжних підодиниць Naa35p і Naa38p. Усі три субодиниці знаходяться на рибосомі у дріжджах, але вони також є у не-рибосомальних формах NAT, як Nat2. NatC-комплекс ацетилює N-кінцевий метіонін субстратів Met-Leu -, Met-Ile-, Met-Trp- або Met-Phe- N-кінців.
NatD
NatD складається лише з каталітичної одиниці Naa40р, яка концептуально відрізняється від інших комплексів NAT. Спочатку було виділено лише два субстрати — H2A та H4 у дріжджів та людини. По-друге, специфічність субстрату Naa40p лежить в межах перших 30-50 залишків, що значно більше, ніж специфічність субстрату інших NAT. Ацетилювання гістонів комплексом NatD частково пов'язане з рибосомами, а амінокислотні субстрати є самими N-кінцевими залишками, що відрізняє його від лізинових N-ацетилтрансфераз (KATs).[12]
NatE
Комплекс NatE складається з субодиниці Naa50p та двох NatA субодиниць: Naa10p і Naa15p. N кінець субстратів Naa50p відрізняється від них NatA ацетильованою активністю Naa10p.[13]
NatF
NatF — це NAT, до складу якого входить фермент Naa60. Спочатку вважалося, що NatF виявлений лише у вищих еукаріотів, оскільки він відсутній у дріжджів.[14] Однак пізніше було встановлено, що Naa60 знаходиться у всьому еукаріотичному домені, хоча вдруге втрачається у лінії грибів.[15] Порівняно з дріжджами, NatF сприяє підвищенню кількості N-кінцевого ацетилювання у людини. NatF-комплекс ацетилює N-кінцевий метіонін субстратів Met-Lys-, Met-Leu-, Met-Ile-, Met-Trp- і Met-Phe- N-закінчення, які частково перекриваються з NatC і NatE.[6] Показано, що NatF локалізується у певних клітинних компартментах та ацетилює цитозольні N-кінці трансмембранних білків.[16] Компартменталізація Naa60 опосередковується його унікальним С-кінцем, який складається з двох альфа-спіралей, які периферично асоціюються з мембраною і опосередковують взаємодію з фосфатидилінозитол-4-фосфатом PI(4)P.[17]
NAA80/NatH
NAA80/NatH — це N-кінцева ацетилтрансфераза, яка специфічно ацетилює N-кінець актину.[18]
Стабілізація білків
N-кінцеве ацетилювання може впливати на стабільність білка, але результати та механізм поки остаточно незрозумілі.[19] Спочатку вважалося, що N-кінцеве ацетилювання захищає білки від деградації, оскільки Nα-ацетилювання N-кінців повинно було блокувати N-кінцеву повсюдність і подальшу деградацію білка.[20] Але ряд досліджень показав, що N-кінцевий ацетильований білок має швидкість деградації подібну до білків із неблокованим N-кінцем.[21]
Локалізація білка
Показано, що N-кінцеве ацетилювання може керувати розташуванням білків. Arl3p — одна з «Arf-подібних» (Arl) GTPаз, яка має вирішальне значення для організації мембранного руху.[22] Nα-ацетилювання Arl3 запущене за допомогою Голджі-мембран-приналежного білка Sys1p спрямовує транспорт вказаного білка до мембранного комплексу апарату Гольджі. Якщо Phe або Tyr замінити на Ala на N-кінці Arl3p, він більше не зможе розташовуватися на мембрані Ґольджі, що вказує на те, що Arl3p потребує своїх природних N-кінцевих залишків, які можуть бути ацетильовані для належної локалізації.[23]
Метаболізм та апоптоз
Також доведено, що білок N-кінцевого ацетилювання пов'язаний з регуляцією клітинного циклу та апоптозу. Генетичне виснаження (нокдаун) NatA або NatC комплексів призводить до індукції р53-залежного апоптозу, що може свідчити про меншу кількість антиапоптотичних білків або зупинку функціонування через зменшення білка N-кінцевого ацетилювання.[24] Проте, на відміну від нього, каспаза-2, ацетильована NatA, може взаємодіяти з адаптерним білком RIP пов'язаним із Ich-1/Ced-3 гомологічними білками зі смертю домен (RAIDD). Це може активувати каспазу-2 і індукує клітинний апоптоз.[25]
Синтез білка
Рибосоми білків відіграють важливу роль в процесі синтезу білка, який може бути ацетильованим на N-кінці. N-кінцеве ацетилювання білків рибосоми може впливати на синтез білка. Зниження на 27 % і 23 % у швидкості синтезу білка спостерігалося з NatA і NatB штамів видалення. Зниження точності трансляції спостерігалася у NatA деформації видалення і дефект в рибосомі був помічений у NatB деформації видалення.[26]
Рак
Запропоновано, що NATи можуть виступати як онко-білки і пухлинні супресори при захворюваннях людей на рак, і експресія NAT може як збільшувати, так і зменшувати ракові клітини. Ектопічна експресія hNaa10p підвищеної проліферації клітин та регуляції генів, що беруть участь в клітинної проліферації та виживання обміну речовин. Надмірна експресія hNaa10p спостерігалася при раку сечового міхура, раку молочної залози та раку шийки матки.[27] Але експресія високого рівня hNaa10p також може пригнічувати ріст пухлини і знижений рівень hNaa10p пов'язаний з поганими прогнозами, великими пухлинами і метастазами у лімфовузлах.
Таблиця 2. Огляд вираження NatA субодиниць в різних ракових тканинах[28]
| Субодиниці Nat | Ракові тканини | Шаблон вираження |
|---|---|---|
| hNaa10 | раку легенів, раку молочної залози, колоректального раку, гепатоцелюлярної карциноми | високо в пухлині |
| hNaa10 | рак легенів, рак молочної залози, рак підшлункової залози, рак яєчників | втрата гетерозиготності в пухлині |
| hNaa10 | рак молочної залози, рак шлунка, рак легенів | високо в первинної пухлини, але низька метастази в лімфовузли. |
| hNaa10 | Немілкоклітинна карцинома легені | низька при пухлинах |
| hNaa15 | папілярний рак щитоподібної залози, рак шлунка | високо в пухлині |
| hNaa15 | нейробластома | високо в просунутій стадії пухлини |
| hNaa11 | гепатоцелюлярна карцинома | втрата гетерозиготності в пухлині |
Ацетилювання і деацетилювання лізину
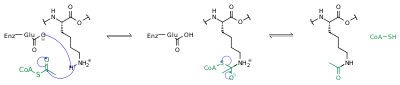
Білки зазвичай ацетильовані на залишках лізину, і ця реакція покладається на ацетил-коензим A як донора ацетильної групи. У процесі ацетилування і деацетилювання гістонів, гістонові білки ацетильовані і деацетильовані на лізинових залишках N-кінцевий хвіст як частина генної регуляції. Зазвичай ці реакції каталізуються ферментами з активністю ацетилтрансферази гістонів (HAT) або гістондеацетилази (деацетилазах гістонів), хоча NATи і HDACи можуть також змінювати статус ацетилювання негістонових білків.[29]
Регуляція факторів транскрипції, ефекторних білків, молекулярних шаперонів та цитоскелетних білків шляхом ацетилювання і деацетилирования є істотним після-трансляційним регуляторним механізмом.[30] Ці регуляторні механізми аналогічні до фосфорилювання і дефосфорилювання під дією кіназ і фосфатаз. Не тільки стан ацетилювання білка може змінювати його активність, але останнім часом припускається, що ця пост-трансляційна модифікація може також надати перехресні переваги для фосфорилювання, метилювання, убіквітинування, сумоїляцією та інші для динамічного контролю клітинної сигналізації.[31] Прикладом цього є регулювання білка тубуліну в нейронах мишей і астроглії.[32][33] Тубулінова ацетилтрансфераза розташована в аксонемі, і ацетилює в α-тубулінову субодиницю в зібраній мікротрубочці. Після того як один раз розібрано, це ацетилювання видаляється іншою специфічною деацетилазою в цитозолі клітини. Таким чином, аксонемальні мікротрубочки, які мають тривалий період напіввиведення, здійснюють «підписне ацетилювання», яке відсутнє у цитозольних мікротрубочках, які мають коротший період напіввиведення.
У галузі епігенетики показано, що ацетилювання гістону (і деацетилювання) є важливими механізмами регуляції транскрипції генів. Однак гістони не є єдиними білками, регульованими посттрансляційним ацетилюванням. Далі наведено приклади різних інших білків, які мають роль у регуляції перетворення сигналу, на діяльність яких також впливає ацетилювання та деацетилювання.
p53
Білок p53 — це пухлинний супресор, який відіграє важливу роль у сигнальних транзакціях у клітинах, особливо у підтримці стабільності геному шляхом запобігання мутації. Тому він також відомий як «охоронець геному». Він також регулює клітинний цикл і затримує ріст клітин, активуючи регулятор клітинного циклу, p21 . При сильному пошкодженні ДНК він також ініціює програму загибелі клітини . Функція p53 негативно регулюється онкобілком Mdm2. Дослідження припускали, що Mdm2 формуватиме комплекс з p53 і не дозволяє йому зв'язуватися з певними генами, чутливими до p53.[34][35]
Ацетилювання р53
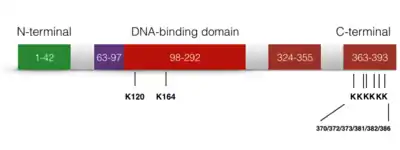
Ацетилювання р53 незамінне для його активації. Повідомлялося, що рівень ацетилювання p53 значно зростає, коли клітина зазнає стресу. На p53 є три основні місця ацетилювання: K164, K120 і C-кінець.[36] Якщо лише одна з ділянок ацетилювання дефектована, активація p21 все ще спостерігається. Однак якщо всі три сайти активації заблоковані, активація p21 та придушення росту клітин, викликане p53, будуть повністю втрачені. Крім того, ацетилювання p53 перешкоджає його зв'язуванню з репресором Mdm2 на ДНК.[37] Також передбачається, що ацетилювання p53 має вирішальне значення для проапоптотичних функцій, незалежних від транскрипції.[38]
Наслідки для терапії раку
Оскільки основна функція p53 — це супресор пухлини, ідея про те, що активація p53 є привабливою стратегією лікування раку. Nutlin-3[39] — це невелика молекула, призначена для націлювання на взаємодію p53 та Mdm2, яка утримувала p53 від дезактивації.[40] Звіти також показали, що ракова клітина під час лікування Nutilin-3a, ацетилювання lys 382, спостерігалася в c-кінці білка p53.[41][42]
Мікротрубочка
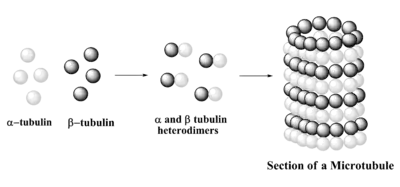
Структура мікротрубочок — це довгий, порожнистий циліндр, який динамічно зібраний із димерів α/β-тубуліну. Вони відіграють важливу роль у підтримці структури клітини, а також клітинних процесах, наприклад, руху органел.[43] Крім того, мікротрубочка відповідає за формування мітотичного веретена в еукаріотичних клітинах для транспортування хромосом при поділі клітин.[44][45]
Ацетилювання тубуліну
Ацетильований залишок α-тубуліну — К40, який у людини каталізується α-тубулін ацетил-трансферазою (α-TAT). Ацетилювання K40 на α-тубулін є ознакою стабільних мікротрубочок. Залишки активної ділянки D157 та C120 α-TAT1 відповідають за каталіз через форму, що доповнює α-тубулін. Крім того, деякі унікальні структурні особливості, такі як шпилька β4-β5, ділянка С-кінцевого циклу та ділянки петлі α1-α2, важливі для специфічного розпізнавання α-тубуліну.[46] Зворотна реакція ацетилювання каталізується гістондеацетилазою 6.[47]
Наслідки для терапії раку
Оскільки мікротрубочки відіграють важливу роль у поділі клітин, особливо у фазі G2/M клітинного циклу, були зроблені спроби унеможливити функціонування мікротрубочок за допомогою інгібіторів малих молекул, які успішно використовуються в клініках як терапія раку.[48] Наприклад, алкалоїди винка та таксани селективно зв'язуються і інгібують мікротрубочки, що призводить до зупинки клітинного циклу.[49] Ідентифікація кристалічної структури ацетилювання α-тубулін ацетил-трансферази (α-TAT) також проливає світло на виявлення невеликої молекули, яка могла б модулювати стабільність або деполімеризацію тубуліну. Іншими словами, орієнтуючись на α-TAT, можна запобігти ацетилюванню тубуліну і призвести до дестабілізації тубуліну, що є аналогічним механізмом дестабілізуючих агентів тубуліну.[46]
STAT3
Перетворювач сигналу та активатор транскрипції 3 (STAT3, від англ. «Signal Transducer and Activator of Transcription 3») — це фактор транскрипції, який фосфорилюється кіназами, пов'язаними з рецепторами, наприклад, тирозинкінази сімейства Януса, і переходить у ядро . STAT3 регулює декілька генів у відповідь на фактори росту та цитокіни та відіграють важливу роль у ріст клітин. Тому STAT3 полегшує онкогенез у різних шляхах, пов'язаних із ростом клітин. З іншого боку, він також відіграє роль в супресорі пухлини .[50]
Ацетилювання STAT3
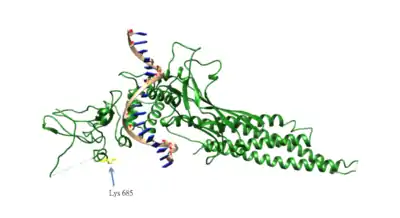
Ацетилювання Lys685 STAT3 має важливе значення для STAT3 гомо-димеризації, що має важливе значення для ДНК-зв'язування та транскрипційної активації онкогенів . Ацетилювання STAT3 каталізується гістон-ацетилтрансферазою p300 і реверсується за допомогою хістондеацетилази типу 1. Ацетилювання лізину STAT3 також підвищено в ракових клітинах.[51]
Терапевтичні наслідки для терапії раку
Оскільки ацетилювання STAT3 важливе для його онкогенної активності та того, що рівень ацетильованого STAT3 високий у ракових клітинах (мається на увазі, що орієнтація на ацетильований STAT3 для хіміопрофілактики та хіміотерапії є перспективною стратегією). Ця стратегія підтримується шляхом лікування ресвератролу, інгібітора ацетилювання STAT3, у раковій клітинній лінії обертається відхилене метилювання острова CpG.[52]
Ацетилювання деревини
З початку 20 століття ацетилювання деревини досліджувалося як метод підвищення міцності деревини в стійкості до процесів гниття та цвілі. До вторинних переваг можна віднести покращення стабільності розмірів, поліпшену твердість поверхні та відсутність зниження механічних властивостей завдяки обробці.
Фізичні властивості будь-якого матеріалу визначаються його хімічною будовою. Деревина містить велику кількість хімічних груп, які називаються «вільні гідроксили». Вільні гідроксильні групи адсорбують і виділяють воду відповідно до змін кліматичних умов, яким піддається деревина. Це пояснення того, чому деревина набухає та усаджується. Вважається також, що перетравлення деревини ферментами ініціюється на вільних ділянках гідроксилу — що є однією з головних причин, чому деревина схильна до гниття.
Ацетилювання змінює вільні гідроксили всередині деревини на ацетильні групи. Це робиться шляхом взаємодії деревини з оцтовим ангідридом, який походить з оцтової кислоти (відомий як оцет, коли він знаходиться в розведеному вигляді). Коли вільна гідроксильна група перетворюється на ацетильну групу, здатність деревини поглинати воду сильно знижується, роблячи деревину стабільною за лінійними параметрами і, оскільки вона вже не піддається дії ферментів, надзвичайно міцна.
У 2007 році лондонська компанія Titan Wood, що має виробничі потужності в Нідерландах, досягла економічної комерціалізації та розпочала масштабне виробництво ацетильованої деревини під торговою назвою «Accoya».
Див. також
- Ацетокси група
- Ациляція
- Амід
- Компендіум ацетилювання білка
- Естер
- Глікозилювання
- Ліпідація
- Нітросилювання
- Органічний синтез
- Протеоліз
Примітки
- Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 325 (5942): 834–840. 2009. Bibcode:2009Sci...325..834C. PMID 19608861. doi:10.1126/science.1175371. Проігноровано невідомий параметр
|vauthors=(довідка) - Mitochondrial acetylome analysis in a mouse model of alcohol-induced liver injury utilizing SIRT3 knockout mice. J. Proteome Res. 11 (3): 1633–1643. 2012. PMC 3324946. PMID 22309199. doi:10.1021/pr2008384. Проігноровано невідомий параметр
|vauthors=(довідка) - Brook, Tom. Protein Acetylation: Much More than Histone Acetylation. Cayman Chemical. Архів оригіналу за 28 лютого 2014.
- Regulation of cellular metabolism by protein lysine acetylation. Science 327 (5968): 1000–1004. 2010. Bibcode:2010Sci...327.1000Z. PMC 3232675. PMID 20167786. doi:10.1126/science.1179689. Проігноровано невідомий параметр
|vauthors=(довідка) - Wang, Qijun; Zhang, Yakun; Yang, Chen; Xiong, Hui; Lin, Yan; Yao, Jun; Li, Hong; Xie, Lu та ін. (19 лютого 2010). Acetylation of Metabolic Enzymes Coordinates Carbon Source Utilization and Metabolic Flux. Science (англ.) 327 (5968): 1004–1007. Bibcode:2010Sci...327.1004W. ISSN 0036-8075. PMC 4183141. PMID 20167787. doi:10.1126/science.1179687.
- NatF contributes to an evolutionary shift in protein N-terminal acetylation and is important for normal chromosome segregation. PLoS Genet. 7 (7): e1002169. 2011. PMC 3131286. PMID 21750686. doi:10.1371/journal.pgen.1002169. Проігноровано невідомий параметр
|vauthors=(довідка) - Protein N-terminal acetyltransferases: when the start matters. Trends Biochem. Sci. 37 (4): 152–161. 2012. PMID 22405572. doi:10.1016/j.tibs.2012.02.003. Проігноровано невідомий параметр
|vauthors=(довідка) - Molecular basis for N-terminal acetylation by the heterodimeric NatA complex. Nat. Struct. Mol. Biol. 20 (9): 1098–105. 2013. PMC 3766382. PMID 23912279. doi:10.1038/nsmb.2636. Проігноровано невідомий параметр
|vauthors=(довідка) - Composition and biological significance of the human Nalpha-terminal acetyltransferases. BMC Proceedings. 3 Suppl 6 (Suppl 6): S3. 2009. PMC 2722096. PMID 19660096. doi:10.1186/1753-6561-3-s6-s3. Проігноровано невідомий параметр
|vauthors=(довідка) - Proteomics analyses reveal the evolutionary conservation and divergence of N-terminal acetyltransferases from yeast and humans. Proc. Natl. Acad. Sci. U.S.A. 106 (20): 8157–8162. 2009. Bibcode:2009PNAS..106.8157A. PMC 2688859. PMID 19420222. doi:10.1073/pnas.0901931106. Проігноровано невідомий параметр
|vauthors=(довідка) - The chaperone-like protein HYPK acts together with NatA in cotranslational N-terminal acetylation and prevention of Huntingtin aggregation. Mol. Cell. Biol. 30 (8): 1898–1909. 2010. PMC 2849469. PMID 20154145. doi:10.1128/mcb.01199-09. Проігноровано невідомий параметр
|vauthors=(довідка) - The human N-alpha-acetyltransferase 40 (hNaa40p/hNatD) is conserved from yeast and N-terminally acetylates histones H2A and H4. PLoS ONE 6 (9): e24713. 2011. Bibcode:2011PLoSO...624713H. PMC 3174195. PMID 21935442. doi:10.1371/journal.pone.0024713. Проігноровано невідомий параметр
|vauthors=(довідка) - The yeast N(alpha)-acetyltransferase NatA is quantitatively anchored to the ribosome and interacts with nascent polypeptides. Mol. Cell. Biol. 23 (20): 7403–7414. 2003. PMC 230319. PMID 14517307. doi:10.1128/mcb.23.20.7403-7414.2003. Проігноровано невідомий параметр
|vauthors=(довідка) - NatF contributes to an evolutionary shift in protein N-terminal acetylation and is important for normal chromosome segregation.. PLoS Genet. 7: e1002169. July 2011. PMC 3131286. PMID 21750686. doi:10.1371/journal.pgen.1002169. Проігноровано невідомий параметр
|vauthors=(довідка) - Rathore, Om Singh; Faustino, Alexandra; Prudêncio, Pedro; Van Damme, Petra; Cox, Cymon J.; Martinho, Rui Gonçalo (2016). Absence of N-terminal acetyltransferase diversification during evolution of eukaryotic organisms.. Scientific Reports 6: 21304. Bibcode:2016NatSR...621304R. PMC 4748286. PMID 26861501. doi:10.1038/srep21304.
- Aksnes (3 березня 2015). An Organellar Nα-Acetyltransferase, Naa60, Acetylates Cytosolic N Termini of Transmembrane Proteins and Maintains Golgi Integrity. Cell Reports 10 (8): 1362–74. PMID 25732826. doi:10.1016/j.celrep.2015.01.053.
- Aksnes, Henriette; Goris, Marianne; Strømland, Øyvind; Drazic, Adrian; Waheed, Qaiser; Reuter, Nathalie; Arnesen, Thomas (2017). Molecular determinants of the N-terminal acetyltransferase Naa60 anchoring to the Golgi membrane.. Journal of Biological Chemistry 292 (16): 6821–6837. PMC 5399128. PMID 28196861. doi:10.1074/jbc.M116.770362.
- NAA80 is actin's N-terminal acetyltransferase and regulates cytoskeleton assembly and cell motility. Proc Natl Acad Sci U S A 115 (17): 4399–4404. 2018. PMC 5924898. PMID 29581253. doi:10.1073/pnas.1718336115. Проігноровано невідомий параметр
|vauthors=(довідка) - N-terminal acetylation and other functions of Nα-acetyltransferases. Biol. Chem. 393 (4): 291–8. 2012. PMID 22718636. doi:10.1515/hsz-2011-0228. Проігноровано невідомий параметр
|vauthors=(довідка) - Role of the alpha-amino group of protein in ubiquitin-mediated protein breakdown. Proc. Natl. Acad. Sci. U.S.A. 81 (22): 7021–5. 1984. Bibcode:1984PNAS...81.7021H. PMC 392068. PMID 6095265. doi:10.1073/pnas.81.22.7021. Проігноровано невідомий параметр
|vauthors=(довідка) - N-terminal acetylation of cellular proteins creates specific degradation signals. Science 327 (5968): 973–977. 2010. Bibcode:2010Sci...327..973H. PMC 4259118. PMID 20110468. doi:10.1126/science.1183147. Проігноровано невідомий параметр
|vauthors=(довідка) - Targeting of the Arf-like GTPase Arl3p to the Golgi requires N-terminal acetylation and the membrane protein Sys1p. Nat. Cell Biol. 6 (5): 405–413. 2004. PMID 15077113. doi:10.1038/ncb1120. Проігноровано невідомий параметр
|vauthors=(довідка) - Knockdown of human N alpha-terminal acetyltransferase complex C leads to p53-dependent apoptosis and aberrant human Arl8b localization. Mol. Cell. Biol. 29 (13): 3569–3581. 2009. PMC 2698767. PMID 19398576. doi:10.1128/mcb.01909-08. Проігноровано невідомий параметр
|vauthors=(довідка) - Depletion of the human Nα-terminal acetyltransferase A induces p53-dependent apoptosis and p53-independent growth inhibition. Int. J. Cancer 127 (12): 2777–2789. 2010. PMID 21351257. doi:10.1002/ijc.25275. Проігноровано невідомий параметр
|vauthors=(довідка) - Metabolic regulation of protein N-alpha-acetylation by Bcl-xL promotes cell survival. Cell 146 (4): 607–620. 2011. PMC 3182480. PMID 21854985. doi:10.1016/j.cell.2011.06.050. Проігноровано невідомий параметр
|vauthors=(довідка) - N(α)-Acetylation of yeast ribosomal proteins and its effect on protein synthesis. J Proteomics 74 (4): 431–441. 2011. PMID 21184851. doi:10.1016/j.jprot.2010.12.007. Проігноровано невідомий параметр
|vauthors=(довідка) - Immunohistochemical analysis of human arrest-defective-1 expressed in cancers in vivo. Oncol. Rep. 21 (4): 909–15. 2009. PMID 19287988. doi:10.3892/or_00000303. Проігноровано невідомий параметр
|vauthors=(довідка) - Protein N-terminal acetyltransferases in cancer. Oncogene 32 (3): 269–276. 2013. PMID 22391571. doi:10.1038/onc.2012.82. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulation of protein turnover by acetyltransferases and deacetylases. Biochimie 90 (2): 306–12. 2008. PMID 17681659. doi:10.1016/j.biochi.2007.06.009. Проігноровано невідомий параметр
|vauthors=(довідка) - Acetylation and deacetylation of non-histone proteins. Gene 363: 15–23. 2005. PMID 16289629. doi:10.1016/j.gene.2005.09.010. Проігноровано невідомий параметр
|vauthors=(довідка) - Lysine acetylation: codified crosstalk with other posttranslational modifications. Mol. Cell 31 (4): 449–61. 2008. PMC 2551738. PMID 18722172. doi:10.1016/j.molcel.2008.07.002. Проігноровано невідомий параметр
|vauthors=(довідка) - Posttranslational modifications of tubulin in cultured mouse brain neurons and astroglia. Biol. Cell 65 (2): 109–117. 1989. PMID 2736326. doi:10.1016/0248-4900(89)90018-x. Проігноровано невідомий параметр
|vauthors=(довідка) - The acetylation of alpha-tubulin and its relationship to the assembly and disassembly of microtubules. J. Cell Biol. 103 (2): 571–579. 1986. PMC 2113826. PMID 3733880. doi:10.1083/jcb.103.2.571. Проігноровано невідомий параметр
|vauthors=(довідка) - Alberts, Bruce (March 2002). Molecular Biology of the Cell. Garland Science. ISBN 0815332181.
- Weinberg, Robert A. (2013). Biology of cancer. (вид. 2.). [S.l.]: Garland Science. ISBN 978-0815342205.
- The impact of acetylation and deacetylation on the p53 pathway. Protein Cell 2 (6): 456–462. 2011. PMC 3690542. PMID 21748595. doi:10.1007/s13238-011-1063-9. Проігноровано невідомий параметр
|vauthors=(довідка) - Acetylation is indispensable for p53 activation. Cell 133 (4): 612–626. 2008. PMC 2914560. PMID 18485870. doi:10.1016/j.cell.2008.03.025. Проігноровано невідомий параметр
|vauthors=(довідка) - p53 acetylation is crucial for its transcription-independent proapoptotic functions. J. Biol. Chem. 284 (17): 11171–11183. 2009. PMC 2670122. PMID 19265193. doi:10.1074/jbc.M809268200. Проігноровано невідомий параметр
|vauthors=(довідка) - In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science 303 (5659): 844–848. 2004. Bibcode:2004Sci...303..844V. PMID 14704432. doi:10.1126/science.1092472. Проігноровано невідомий параметр
|vauthors=(довідка) - Small-molecule inhibitors of the MDM2-p53 protein-protein interaction to reactivate p53 function: a novel approach for cancer therapy. Annu. Rev. Pharmacol. Toxicol. 49 (1): 223–241. 2009. PMC 2676449. PMID 18834305. doi:10.1146/annurev.pharmtox.48.113006.094723. Проігноровано невідомий параметр
|vauthors=(довідка) - Nutlin-3a, an MDM2 antagonist and p53 activator, helps to preserve the replicative potential of cancer cells treated with a genotoxic dose of resveratrol. Mol. Biol. Rep. 40 (8): 5013–5026. 2013. PMC 3723979. PMID 23666059. doi:10.1007/s11033-013-2602-7. Проігноровано невідомий параметр
|vauthors=(довідка) - Nutlin-3a activates p53 to both down-regulate inhibitor of growth 2 and up-regulate mir-34a, mir-34b, and mir-34c expression, and induce senescence. Cancer Res. 68 (9): 3193–3203. 2008. PMC 2440635. PMID 18451145. doi:10.1158/0008-5472.CAN-07-2780. Проігноровано невідомий параметр
|vauthors=(довідка) - Kreis, ed. by Thomas; Vale, Ronald (1999). Guidebook to the cytoskeletal and motor proteins (вид. 2.). Oxford [u.a.]: Oxford Univ. Press. ISBN 0198599560.
- al.], Harvey Lodish ... [et (2013). Molecular cell biology (вид. 7th). New York: W.H. Freeman and Co. ISBN 978-1429234139.
- Fojo, edited by Tito (2008). The role of microtubules in cell biology, neurobiology, and oncology (вид. [Online-Ausg.]). Totowa, N. J.: Humana Press. ISBN 978-1588292940.
- Structure of the α-tubulin acetyltransferase, αTAT1, and implications for tubulin-specific acetylation. Proc. Natl. Acad. Sci. U.S.A. 109 (48): 19655–19660. 2012. Bibcode:2012PNAS..10919655F. PMC 3511727. PMID 23071314. doi:10.1073/pnas.1209357109. Проігноровано невідомий параметр
|vauthors=(довідка) - HDAC6 is a microtubule-associated deacetylase. Nature 417 (6887): 455–458. 2002. Bibcode:2002Natur.417..455H. PMID 12024216. doi:10.1038/417455a. Проігноровано невідомий параметр
|vauthors=(довідка) - Teresa Carlomagno, ред. (2009). Tubulin-binding agents : synthetic, structural, and mechanistic insights. Berlin: Springer. ISBN 978-3540690368.
- Zito, edited by Thomas L. Lemke, David A. Williams; associate editors, Victoria F. Roche, S. William (2013). Foye's principles of medicinal chemistry (вид. 7th). Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins. ISBN 978-1609133450.
- Müller-Decker, Friedrich Marks, Ursula Klingmüller, Karin (2009). Cellular signal processing : an introduction to the molecular mechanisms of signal transduction. New York: Garland Science. ISBN 978-0815342151.
- Stat3 dimerization regulated by reversible acetylation of a single lysine residue. Science 307 (5707): 269–273. 2005. Bibcode:2005Sci...307..269Y. PMID 15653507. doi:10.1126/science.1105166. Проігноровано невідомий параметр
|vauthors=(довідка) - Acetylated STAT3 is crucial for methylation of tumor-suppressor gene promoters and inhibition by resveratrol results in demethylation. Proc. Natl. Acad. Sci. U.S.A. 109 (20): 7765–7769. 2012. Bibcode:2012PNAS..109.7765L. PMC 3356652. PMID 22547799. doi:10.1073/pnas.1205132109. Проігноровано невідомий параметр
|vauthors=(довідка)
