Фактори транскрипції
Факторами транскрипції (інша назва — специфічні до послідовності ДНК-зв'язувальні фактори) в молекулярній біології називають білки, що зв'язуються із регуляторними ділянками ДНК за допомогою своїх ДНК-зв'язувальних доменів і є частиною системи, яка регулює транскрипцію, тобто передачу генетичної інформації від ДНК до РНК[1][2].
Різні фактори транскрипції можуть як сприяти зв'язуванню РНК-полімерази з промотором (у такому випадку спостерігається активація транскрипції, а сам фактор називається «активатором»), так і запобігати зв'язуванню РНК-полімерази (у такому випадку відбувається репресія транскрипції, а сам фактор називається «репресором»)[3][4]. Фактори транскрипції виконують таку функцію або самостійно, або використовуючи інші допоміжні білки. Залежно від функції, ці білки також поділяються на «коактиватори» та «корепресори».
Функція
Базальні фактори транскрипції
В еукаріотів важливий клас факторів транскрипції, необхідних для будь-якої транскрипції, називається базальними факторами транскрипції[5][6] Деякі з них не зв'язуються безпосередньо з ДНК, але є частиною великого комплексу ініціації разом з РНК-полімеразою. До базальних факторів транскрипції належать TFIIA, TFIIB, TFIID, TFIIE, TFIIF, and TFIIH.[7] Приєднуючись до ДНК в районі промотору, вони створюють «платформу» для РНК-полімерази, на яку вона сідає, і таким чином ініціюють транскрипцію. Базальні фактори транскрипції відрізняються за своєю структурою і функціями. Одні містять регіони для зв'язку з ТАТА-боксом на ДНК (ТАТА-сполучний протеїн транскрипційного фактору TFIID), інші мають протеїнкіназну або геліказну активність, наприклад, TAF250-TFIID. Базальні фактори транскрипції убіквітарні (ubiquitarius; лат. ubique всюди), тобто присутні рівномірно в усіх клітинах всього організму і на специфічну експресію генів не впливають.
Специфічні фактори транскрипції
Специфічні фактори транскрипції «повідомляють» РНК-полімеразі, який саме ген необхідно активувати. Вони знаходяться тільки в тих клітинах, в яких вони мають активувати (або репресувати) певний ген. Промотори, з якими вони зв'язуються, мають певні характерні нуклеотидні послідовності (енхансери або сайленсери), які розпізнають фактори транскрипції і сполучаються з ними. Специфічні фактори транскрипції часто активуються протеїнкіназами, і така активація є останньо ланкою тривалого процесу передачі клітинного сигналу, що розпочався з активації рецептору.
Процеси, що регулюються факторами транскрипції
Розвиток
У дріжджів за певних умов культивування може спостерігатись морфологічний розвиток у волокнисту структуру. Цей розвиток регулюють фактори транскрипції Ste12 та Tec1[8].
У рослин фактори транскрипції регулюють розвиток і формування насіння, його проростання, ріст проростка, формування таких органів з меристематичної тканини, як пагін та квітка. Прикладом регуляції морфогенезу факторами транскрипції у покритонасінних може бути ABC модель розвитку квітки. Гени, які контролюють ідентичність меристеми суцвіття належать до MADS сімейтва факторів транскрипції. Гени класу А (APETALA2 (AP2) та APETALA1 (AP1)) контролюють розвиток чашолистиків і пелюсток. Гени класу В (APETALA3 (AP3) та PISTILLATA (PI)) відповідають за пелюстки і тичники, а гени класу С (AGAMOUS (AG)) за розвиток тичинок і гінецію[9].
Численні фактори транскрипції задіяно в регуляції розвитку багатоклітинних організмів — онтогенезі. У відповідь на отриманий сигнал такі фактори транскрипції вмикають або вимикають транскрипцію генів, що призводить до зміни морфології клітини і визначає її диференціацію. Наприклад, фактор транскриції з сімейства Hox відповідає за належну схему формування тіла, починаючи з плодової мухи дрозофіли, закінчуючи людиною[10][11]. Іншим прикладом фактору транскрипції, задіяного в ембріогенезі, є SRY, який відіграє вирішальну роль в формуванні статі людського плоду[12].
Клітинний цикл
Багато транскрипційних факторів задіяно в регуляції клітинного циклу. Вони визначають, до якого розміру має вирости клітина, коли саме вона має вступити в процес поділу і коли завершити такий процес. Прикладом такого транскрипційного фактору у дріжджів є Swi5, робота якого приводить до деактивації специфічної протеїнкінази і, як наслідок, виходу клітин зі стадії мітотичного поділу[13].
У рослин клітинний цикл, а саме консерватнивний регуляторний шлях переходу від G1(G0) до S фази, регулює фактор транскрипції E2F. Подібний за структурою фактор транскрипції існує й у тварин. Він відіграє схожу функцію у контролі клітинного поділу[14]. Іншим прикладом регуляції клітинного циклу у тварин є Myc-онкоген, який відіграє роль в рості клітини і апоптозі.
Відповідь на внутрішньоклітинні сигнали
Клітини можуть комунікувати між собою шляхом виділення в позаклітинне середовище молекул, які сприймає система рецепторів іншої клітини. Отриманий сигнал запускає каскад передачі молекулярного сигналу всередині клітини, що передається до факторів транскрипції і, як результат, приводить до ввімкнення або вимикнення генів, що їх регулює відповідний фактор транскрипції.
Прикладом подібної комунікації у рослин може слугувати регуляція експресії генів під дією етилену під контролем етилен-чутливих факторів транскрипції. Етилен є фітогормоном, синтез якого відбувається в клітинах рослин і який регулює дозрівання фруктів та «старіння» й опадання листя восени[15].
У тварин прикладом подібної регуляції виступає естроген. Секретований тканинами яєчників і плаценти, естроген проникає крізь клітинну мембрану клітин-реципієнтів, зв'язується з естрогеновим рецептором у цитоплазмі і транспортується в клітинне ядро, де прикріплюється до характерних ділянок ДНК у промоторному регіоні. Наслідком чого є зміна транскрипційної активності відповідних генів.
Відповідь на умови зовнішнього середовища
Будь-яку зміну умов навколишнього середовища вловлюють рецепторні системи клітини. Специфічний сигнал проходить до факторів транскрипції, що має наслідком активацію генів, продукти яких задіяно в адаптації до змінених умов — змін у температурі чи вологості, рівні кисню, кислотності, ультрафіолетового випромінюванні чи освітлюванності, наявності поживних речовин.
Наприклад, у дріжджів при культивування на середовищі з достатньою кількістю моно- і дисахаридів виключені всі гени, задіяні в метаболізмі альтернативних поживних речовин, таких як рафіноза. Таке явище має назву глюкозна репресія. За відсутності легкодоступних вуглеводів системи глюкозної репресії вимикаються, і фактори транскрипції вмикають гени, відповідальні за метаболізм альтернативних джерел вуглеводів.
У тварин транскрипційний фактор теплового шоку HSF (Heat Shock Factor) вмикає експресію генів, продукти яких допомагають клітині вижити в умовах підвищеної температури, а фактор, індукований гіпоксією (HIF, Hypoxia-Inducible Factor), вмикає гени, що захищають клітину в умовах кисневого голоду.
Регуляція
Зазвичай більшості біологічних процесів притаманні багаторівнева регуляція і контроль. Це стосується й факторів транскрипції — вони не тільки самостійно контролюють ефективність транскрипції в регулюванні кількості будь-якого генного продукту (мРНК та білка) в клітині, наявність факторів транскрипції й ефективність їхньої роботи сама складно регулюється. Нижче наведено короткий огляд цих регуляторних шляхів.
Синтез
Як будь-який білок, фактор транскрипції закодовано у вигляді гену, що зчитується з хромосоми у вигляді мРНК, яка згодом транслюється у білок. Кожен з таких етапів синтезу може регулюватись, що впливає на виробництво і, відповідно, активність фактору транскрипції. Цікавою особливістю регуляції на даному етапі є те, що фактор транскрипції може регулювати сам себе за допомогою негативного зворотного зв'язку. В цьому випадку він може діяти як власний репресор: якщо фактор транскрипції приєднується до промотору перед власним геном, це призводить до зменшення синтезу РНК зі свого власного гену[16]. Це один з механізмів, що пояснює загальний низький рівень експресії факторів транскрипції у клітині. У деяких випадках зворотний зв'язок може бути й позитивним, що приводить до різкої відмінності у кількості (або рівні активності) фактора транскрипції на різних стадіях життя клітини[17].
Нещодавно відкриті мікроРНК відіграють суттєву роль в регуляції транскрипції генів шляхом репресії трансляції білка з мРНК або сприянню деградації мРНК, комплементарно зв'язуючись з мішенню. В багатьох випадках мРНК факторів транскрипції якраз і виступає безпосередньою мішенню деяких мікроРНК. Наприклад, у рослин експресія ТСР генів, що кодують фактори транскрипції регулює мікроРНК[18].
Транспорт
В еукаріотів гени факторів транскрипції, як і більшість білків, транскрибуються в клітинному ядрі, після чого їхні мРНК транспортуються у цитоплазму, де і проходить процес трансляції. Втім, білки, що обслуговують ядерні процеси, повертаються назад до ядра в процесі ядерного транспорту. Сигналом для цього служить наявність в білковій послідовності спеціального регіону, який називається сигнальною послідовністю ядерної локалізації. Ця послідовність грає значну роль в регуляції факторів транскрипції.[19]. Деякі фактори транскрипції, наприклад, певні ядерні рецептори та деякі інші, перш ніж їх буде транспортовано до ядра, також повинні приєднати певний необхідний для їхньої активації ліганд ще у цитоплазмі.
Активація
Деякі фактори транскрипції мають у своїй структурі крім домену, за допомогою якого фактор прив'язується до ДНК, ще цілу низку регуляторних, чутливих до певних молекулярних сигналів доменів. За механізмом сприйняття сигналу їх умовно можна поділити на такі категорії:
- зв'язок з лігандом. Як було зазначено вище, зв'язок з лігандом може визначати ефективність або можливість ядерної локалізації фактору. Так само зв'язок з лігандом може змінювати просторову структуру білка і таким чином впливати на ефективність взаємодії білок-ДНК, а також на білок-білкову взаємодію.
- фосфорилювання. Багато факторів транскрипції, наприклад STAT, у своїй структурі містять специфічні ділянки, які повинні фосфорилюватись, перші ніж фактор транскрипції зв'яжеться з ДНК[20][21].
- взаємодія з іншими факторами транскрипції. Часто фактори транскрипції здатні до гомо-, гетеро- і димеризації або утворення білкових комплексів з регуляторними білками.
Фізична доступність ділянок ДНК до зв'язування з фактором
В еукаріотів гени, які не транскрибуються на даний момент, часто локалізуються в гетерохроматині. Гетерохроматин — це щільно упакована разом з гістонами ділянка хромосоми. ДНК в упакованому вигляді фізично недоступна для більшості факторів транскрипції. Для того, щоб фактор транскрипції мав можливість приєднатись до ДНК, гетерохроматин повинен розкрутитись в еухроматин — менш щільну організацію хромосоми. Це здійснюється шляхом модифікації гістонів, наприклад, їхнім метилюванням. Таким чином регуляція структури хроматину опосередковано впливає на регуляцію роботи факторів транскрипції. Іншим чинником фізичної недоступності може бути зайнятість ділянки ДНК іншим фактором транскрипції. Фактори транскрипції можуть працювати в парі, граючи антагоністичні ролі (активатор — репресор) в регуляції транскрипції одного і того самого гену.
Наявність інших кофакторів і факторів транскрипції
Більшість факторів транскрипції не працюють самостійно. Початок процесу транскрипції вимагає одночасної роботи багатьох білків. Відсутність бодай одного може призвести до неефективної роботи всього комплексу ініціації і РНК-полімерази.
Структура
Фактори транскрипції мають модульну структуру і містять такі домени:
- ДНК-зв'язувальний домен, за допомогою якого фактор транскрипції безпосередньо зв'язуюється з ДНК у районі промоторів.
- Домен транс-активації, що містить ділянки, куди можуть приєднуватись інші білки, такі як транскрипційні ко-регулятори. Ці домени часто фігурують в контексті активаторної функції[22]
- Домен, чутливий до сигналу, або, як його ще називають, ліганд-зв'язувальний домен, який може розпізнавати зовнішній молекулярний сигнал і відповідно змінювати активність фактору транскрипції, що приводить до загальної зміни в експресії генів. Чутливість до сигналу і функція трансактивації може бути як в одному і тому самому домені, так і знаходитись не тільки в різних доменах, а й в різних білках.
Домен ДНК-прив'язки
Домен ДНК-прив'язки — будь-який білковий мотив, який зв'язується з подвійним або одинарним ланцюгом ДНК завдяки спорідненості до ДНК взагалі або до конкретної специфічної послідовності нуклеотидів. Нижче наведено список декількох основних доменів ДНК-прив'язки.
| Родина | Структурна класифікація білків SCOP | База даних InterPro |
|---|---|---|
| основна спіраль-петля-спіраль basic helix-loop-helix (bHLH)[23] | SCOP 47460 | IPR001092 |
| основний лейциновий zipper basic leucine zipper (bZIP)[24] | SCOP 57959 | IPR004827 |
| C-термінальний ефекторний домен регуляторів двосторонньої відповіді | SCOP 46894 | IPR001789 |
| GCC бокс | SCOP 54175 | |
| спіраль-поворот-спіраль helix-turn-helix (HTH)[25] | ||
| гомеодоменний протеїн — зв'язуюється з промоторною послідовність генів з сімейства, які мають гомеобокс послідовність і які самі є факторами транскрипції, що задіяні в регуляції розвитку .[26] | SCOP 46689 | IPR009057 |
| парний бокс, який містять гени сімейства pax[27] | ||
| крилата спіраль | SCOP 46785 | IPR011991 |
| цинкові пальці[28] | ||
| * мульті-домен Cys2His2 цинкові-пальці[29] | SCOP 57667 | IPR007087 |
| * Zn2/Cys6 | SCOP 57701 | |
| * Zn2/Cys8 ядерний рецептор цинкові пальці | SCOP 57716 | IPR001628 |
Ділянка зв'язування
Послідовність ДНК, до якої прикріплюється фактор транскрипції називається ділянкою зв'язування. Фактори транскрипції взаємодіють хімічно з цією ділянкою за допомоги комбінації електростатичних або ван дер Ваальсових сил. Природа цих хімічних взаємодій визначає специфічність приєднання до ДНК більшості факторів транскрипції, проте сама сила відповідної взаємодії залежить від нуклеотидної послідовності. Деякі фактори транскрипції здатні приєднуватись не до однієї послідовності, а до різних, якщо вони подібні, утворюючи різні за силою зв'язки.
Наприклад, ТАТА-сполучний протеїн специфічно розпізнає нуклеотидну послідовність TATAAAA (TATA-бокс), але здатен також приєднуватись до таких послідовностей, як TATATAT або TATATAA.
Оскільки фактори транскрипції можуть приєднуватись до цілої низки нуклеотидних послідовностей, які ж до того достатньо короткі і зустрічаються часто на всьому геномі, малоймовірно, що фактори транскрипції приєднаються до будь-якої подібної послідовності на будь-якій ділянці ДНК. Ймовірно, що фізична доступність ділянки ДНК, наявність кофакторів сприяють визначенню, який саме фактор транскрипції буде працювати на цій конкретній ділянці ДНК. Сама нуклеотидна послідовність не є достатньою інформацією для передбачення потенційної ділянки зв'язування.
Втім, існують комп'ютерні програми, які можуть передбачити можливі ділянки зв'язування завдяки алгоритму порівнянню нуклеотидних послідовностей з уже відомими і описаними раніше, як наприклад TFSEARCH[30], AliBaba 2.1, F-Match 1.0, Match — 1.0[31].
Класифікація факторів транскрипції
Оскільки фактори транскрипції мають багато подібностей у побудові доменів, хоча механізми регуляції, а також процеси, які вони регулюють, суттєво відрізняються, існує три незалежних класифікації факторів транскрипції: (1) класифікація за механізмом дії (2) функціональна класифікація та (3) класифікація за подібністю структурних доменів. Всі три типи класифікації стосуються одночасно всіх типів живих організмів.
Класифікація за механізмом дії
Розрізняють три основних типи за механізмом дії:
- Базальні фактори транскрипції, задіяні у формуванні комплексу преініціації. Як було згадано вище, до таких належать фактори TFIIA, TFIIB, TFIID, TFIIE, TFIIF і TFIIH, які є убіквітарними і зв'язуються з ДНК у регіоні навколо транскрипційного старту генів класу II.[32]
- Верхні фактори транскрипції - фактори, які приєднуються до промотору вище послідовності ініціації транскрипції, стимулюючи або репресуючи транскрипцію.
- Транскрипційні фактори, які індукуються — фактори, які подібні до попередньої групи, але вимагають певного молекулярного сигналу для приєднання до транскрипційного комплексу.
Функціональна класифікація
Фактори транскрипції класифікують за їхньої регуляторною функцією:
- I. конститутивні — присутні в усіх клітинах в будь-який момент часу. Наприклад, базальні фактори транскрипції, Sp1, NF1, Ccaat-енхансер-зв'язувальний протеїн.
- II. індуцибельні — такі, що вимагають активації..
- II.A залежні від процесу розвитку (клітинно-специфічні) — їхня експресія контрольована, втім, якщо вони вже наявні в клітині, то не вимагають додаткової активації — GATA, HNF, PIT-1, MyoD, Myf5, Hox.
- II.B залежні від сигналу — такі, що вимагають зовнішнього сигналу для активації.
- II.B.1 залежні від зовнішньоклітинного сигналу-ліганду .
- II.B.2 залежні від внутрішньоклітинного сигналу-ліганду — SREBP, p53.
- II.B.3 залежні від рецепторів клітинної мембрани.
- II.B.3.a резидентні ядерні фактори — такі, що знаходяться в ядрі в залежності від отриманого сигналу. Наприклад: CREB, AP-1, Mef2.
- II.B.3.b латентні цитоплазматичні фактори — неактивна форма фактору транскрипції, що міститься в цитоплазмі і при отриманні відповідного сигналу й активації прямує до ядра. Наприклад: STAT, R-SMAD, NF-kB, Notch, TUBBY, NFAT.
Класифікація за подібністю структурних доменів
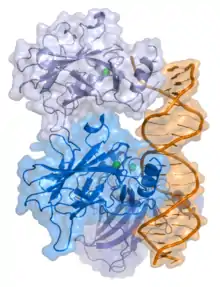
Найчастіше фактори транскрипції класифікують за подібністю до функціональних доменів і третинної структури їхнього домену ДНК-прив'язки[33][34][35].
- 1 Надклас: Основні Домени (основна спіраль-петля-спіраль) basic helix-loop-helix (bHLH)
- 1.1 Клас: Лейцинова застібка (Leucine zipper bZIP)
- 1.1.1 Родина: AP-1 -подібні
- 1.1.2 Родина: CREB
- 1.1.3 Родина: Ccaat-енхансер-зв'язувальний протеїн-подібні
- 1.1.4 Родина: bZIP / PAR
- 1.1.5 Родина: Рослинні G-box зв'язувальні фактори
- 1.1.6 Родина: ZIP тільки
- 1.2 Клас: Спіраль-петля-спіраль
- 1.2.1 Родина: Убіквітарні фактори клас А
- 1.2.2 Родина: Міогенні фактори MyoD
- 1.2.3 Родина: Achaete-Scute
- 1.2.4 Родина: Tal/Twist/Atonal/Hen
- 1.3 Клас: Спіраль-петля-спіраль / Лейцинова застібка bHLH-ZIP
- 1.3.1 Родина: Убіквітарні bHLH-ZIP фактори, включаючи USF та SREBP
- 1.3.2 Родина: Фактори, які контролюють клітинний цикл c-Myc
- 1.4 Клас: NF-1
- 1.4.1 Родина: NF-1
- 1.5 Клас: RF-X
- 1.5.1 Родина: RF-X (гени NFX2, NFX3, NFX5)
- 1.6 Клас: bHSH
- 1.1 Клас: Лейцинова застібка (Leucine zipper bZIP)
- 2 Надклас: Цинк-координовані домени ДНК-прив'язки
- 2.1 Клас: Cys4 цинкові пальці (zinc finger) ядерних рецепторів
- 2.1.1 Родина: Рецептори стероїдних гормонів
- 2.1.2 Родина: Подібні до рецепторів тироїдних гормонів
- 2.2 Клас: різноманітні Cys4 цинкові пальці
- 2.2.1 Родина: GATA фактори транскрипції
- 2.3 Клас: Cys2His2 домен цинкові пальці
- 2.3.1 Родина: Убіквітарні фактори TFIIIA та Sp1
- 2.3.2 Родина: Регулятори розвитку і клітинного циклу Krüppel
- 2.3.4 Родина: Великі фактори зі зв'язувальними якостями / NF-6B-подібні
- 2.4 Клас: Cys6 цистеїн-цинковий кластер
- 2.5 Клас: Цинкові пальці іншої композиції
- 2.1 Клас: Cys4 цинкові пальці (zinc finger) ядерних рецепторів
- 3 Надклас: Спіраль-поворот-спіраль
- 3.1 Клас: Гомеобокс
- 3.1.1 Родина: Тільки гомеодомен Ubx
- 3.1.2 Родина: POU домен
- 3.1.3 Родина: Гомеодомен з регіоном LIM
- 3.1.4 Родина: Гомеодомен плюс цинкові пальці
- 3.2 Клас: Парний бокс
- 3.2.1 Родина: Парний бокс плюс гомеодомен
- 3.2.2 Родина: Тільки парний бокс
- 3.3 Клас: Вилкова головка (Fork head) / Крилата спіраль (Winged helix)
- 3.3.1 Родина: Регулятори розвитку forkhead
- 3.3.2 Родина: Тканинно-специфічні регулятори
- 3.3.3 Родина: Регулятори клітинного циклу
- 3.3.0 Родина: Інші регулятори
- 3.4 Клас: Фактори теплового шоку HSF
- 3.4.1 Родина: HSF
- 3.5 Клас: Триптофановий кластер
- 3.5.1 Родина: Myb
- 3.5.2 Родина: Ets-подібний
- 3.5.3 Родина: Інтерферон-регульований
- 3.6 Клас: TEA домен транскрипційний енхансер
- 3.1 Клас: Гомеобокс
- 4 Надклас: бета-складчасті фактори з незначним жолобковим контактом
- 4.1 Клас: RHR
- 4.1.1 Родина: Rel/ankyrin, NF-kB
- 4.1.2 Родина: ankyrin тільки
- 4.1.3 Родина: NF-AT
- 4.2 Клас: STAT
- 4.2.1 Родина: STAT
- 4.3 Клас: p53
- 4.3.1 Родина: p53
- 4.4 Клас: MADS бокс
- 4.4.1 Родина: регулятори диференціації
- 4.4.2 Родина: ті, що відповідають на зовнішній сигнал
- 4.4.1 Родина: регулятори диференціації
- 4.5 Клас: бета-Barrel альфа-спіраль
- 4.6 Клас: TATA-зв'язувальний протеїн
- 4.8 Клас: Гетеромерні CCAAT фактори
- 4.8.1 Родина: Гетеромерні CCAAT фактори
- 4.9 Клас: Grainyhead
- 4.9.1 Родина: Grainyhead
- 4.10 Клас: Домен холодового шоку
- 4.10.1 Родина: csd
- 4.11 Клас: Runt
- 4.11.1 Родина: Runt
- 4.1 Клас: RHR
- 0 Надклас: Інша фактори транскрипції
- 0.1 Клас: Copper перший протеїн
- 0.2 Клас: HMGI(Y)
- 0.2.1 Родина: HMGI(Y)
- 0.3 Клас: Pocket домен
- 0.4 Клас: E1A-подібний
- 0.5 Клас: AP2/EREBP-подібний
- 0.5.1 Родина: Apetala 2
- 0.5.2 Родина: EREBP
- 0.5.3 Надродина: AP2/B3
- 0.5.3.1 Родина: ARF
- 0.5.3.2 Родина: ABI
- 0.5.3.3 Родина: RAV
Методи дослідження факторів транскрипції
Комп'ютерні методи дослідження
Методи передбачення ділянок зв'язування для факторів транскрипції
- Метод дослідження нуклеотидних послідовностей.
Наразі найпоширеніший метод передбачення потенційних ділянок зв'язування, який базується на порівняння з уже відомими раніше описаними ділянками. Для пошуку використовують алгоритм матриць позиційної ваги (Position Weight Matrix, PWM)[36] Втім, точність передбачення залежить від позиції ділянки на хромосомі і якості прочитання ділянки ДНК.
- Метод ab-initio (лат. від початку).
Цей метод не базується на експериментальних даних, а радше на компьютерній симуляції контакту нуклеотидів і амінокислот. Комп'ютерні моделювання, які розглядають структурну гнучкість і надмірність взаємодії, вимагають інтенсивного і тривалого обчислення. «Карти вільної-енергії», отримані від обчислень взаємодії різних пар нуклеотидів і амінокислот, показали різну специфічність. Ці дані для усіх комбінацій нуклеотидів і амінокислот можуть бути використані для передбачення ділянок зв'язування.[37]
- Метод порівняння структур.
Полягає в аналізі бази даних структур комплексу ДНК — білок. В цьому випадку потенційні функції для специфічних взаємодій між нуклеотидами і амінокислотами визначаються емпірично, як результат статистичного аналізу. Для оцінки здатності послідовностей до складних структур специфічних факторів транскрипції використовують статистичний потенціал і комбінацію послідовних обчислювальних процедур, подібну до розпізнавання тримірної структури протеїну. Точність цього методу для цільового передбачення все ще обмежена через обмежену кількість доступних структурних даних. Проте, перевага цього методу полягає в тому, що можна кількісно дослідити специфічність ефектів деформації ДНК та інші структурні ефекти. Потенційне збільшення кількості структурних даних робить цей метод доволі перспективним.[38]
Лабораторні методи дослідження
Пошук нових факторів транскрипції.
Як і будь-який ген, ген, що кодує фактор транскрипції, а також промоторну послідовність ДНК, можна клонувати, використовуючи стандартні технології та інформацію, отриману за допомогою комп'ютерних методів дослідження. Клонований ген можна зберігати у векторі і використовувати для різноманітних експериментів — від детекції гену в геномі до генно-інженерних маніпуляцій.
- ChIP-on-chip
Новий потужний експериментальний метод, який є комбінацією імунопреципітації хроматину (chromatin immunoprecipitation («ChIP»))і технології біочипа. Метод дозволяє вивчати взаємодію білків і ДНК in vivo.[39] Найчастіше його використовують в експериментах з організмами, геном яких прочитано повністю, що дає максимальну кількість інформації щодо нуклеотидних послідовностей промоторів. До цих послідовностй приєднуються білки, які можуть бути потенційними факторами транскрипції, завдяки здатності приєднуватись фізично до ДНК. Ці білки можна детектувати, виділяти і секвенувати. Недоліками цього методу є висока ціна, великий розмір ДНК фрагментів, що аналізуть, а також неможливість розрізняти геномні повтори.
Вивчення структури і фізико-хімічних властивостей факторів транскрипції
- Метод оцінки термодинамічного потенціалу, або енергії Гіббса (дельта G).
Цей метод базується на експериментальному вимірюванні хімічної спорідненості фактору транскрипції до ділянки зв'язування. Оскільки в даному випадку вимірюється фізична можливість зв'язку, передбачення є достовірнішим. Метод має певні обмеження, оскільки не враховує можливої участі інших факторів транскрипції для зв'язування, а також зміни тривимірної конфігурації молекули при отриманні додаткових молекулярних сигналів.[40]
Вивчення функції факторів транскрипції.
- Полімеразна ланцюгова реакція (ПЛР) у реальному часі.
Реакція зворотної транскрипції в комбінації з ПЛР в реальному часі дає змогу кількісно оцінювати рівень експресії генів, що кодують фактори транскрипції. Метод дозволяє одночасно досліджувати експресію всіх сьогодні відомих факторів транскрипції у різних тканинах. Наприклад для арабідопсису розроблена платформа ПЛР в реальному часі, де аналізу піддають одночасно 1400 факторів транскрипції[41]
Біочип також широко використовується для вивчення експресії генів у різних тканинах. Як і для ПЛР в реальному часі, мРНК повинна бути зворотно-транскрибована в кДНК, помічена радіоактивною або флюоресцентною міткою і загібридизована на біочип. Метод дає можливість порівнювати мРНК з різних тканин.[42]
- Візуалізація роботи факторів транскрипції in vivo.
Відкриття зеленого флюоресцентного білка і застосування його в лабораторних дослідженнях відкрило нові можливості для візуалізації локалізації і динаміки міграції факторів транскрипції в клітині. Для цього використовують комбінацію генно-інженерних підходів і флюоресцентну мікроскопію.[43]
Значення для людини
Хвороби
Оскільки фактори транскрипції регулюють експресію цілої низки генів, мутації у них, як правило, значно впливають на фенотипові прояви, що стосуються процесів розвитку і клітинного циклу.
У людини описано кілька хвороб, викликаних мутацією в генах, що кодують фактори транскрипції.
- Синдром Ретта. Психоневрологічна хвороба, викликана мутацією в транскрипційному факторі MECP2[44]. Це репресор, який працює в певний момент часу в мозку дитини і є відповідальним за вимкнення у певний момент кількох генів протягом розвитку мозку. Якщо ген мутований, то виключення не відбувається і мозок починає неправильно розвиватись. Ген локалізований на Х-хромосомі, тому хвороба спостерігається тільки у дівчаток. Для плоду чоловічої статі ця мутація летальна.
- Діабет. Одна з рідких форм діабету може бути викликана мутаціями в генах факторів транскрипції, один з яких називається гепатоцитний ядерний фактор HNF[45], а інший — фактор інсулінового промотору IPF1[46].
- Вербальна апраксія. Хвороба, коли пацієнт не здатний до координованих рухів і жестів, які супроводжують мову. Викликана мутацією в факторі транскрипції FOXP2[47].
Одомашнення та селекція
Порівняльний аналіз геномів рослин показав, що гени, які кодують фактори транскрипції, еволюціонують найшвидше. Причому саме на добір мутацій цих генів була направлена штучна селекція рослин, яка привела до їхнього одомашнення.
- Одомашнення кукурудзи. Мутація гену, що кодує фактор транскрипції tga1, який належить до сімейства SBP-доменів призвела до перетворення предка кукурудзи теосінте на сучасну кукурудзу[50].
- Одомашнення томатів. Згідно з результатами наукового дослідження, розмір фруктів томатів зумовлений мутацією в факторі транскрипції, який гомологічний до людського онкогену c-H-ras p21[51].
- Одомашнення пшениці. Основним геном, мутація в якому посприяла утворенню одомашненої форми пшениці, був ген Q, який належить до факторів транскрипції сімейства AP2. Цей ген контролює цілий ряд фенотипових ознак, такі як форму і пружність зернівки, довжину колоска, крихкість стебла, а також час колосіння[52].
Посилання
- Latchman DS (1997). Transcription factors: an overview. Int. J. Biochem. Cell Biol. 29 (12): 1305–12. PMID 9570129. doi:10.1016/S1357-2725(97)00085-X.
- Karin M (1990). Too many transcription factors: positive and negative interactions. New Biol. 2 (2): 126–31. PMID 2128034.
- Roeder RG (1996). The role of general initiation factors in transcription by RNA polymerase II. Trends Biochem. Sci. 21 (9): 327–35. PMID 8870495. doi:10.1016/0968-0004(96)10050-5.
- Nikolov DB, Burley SK (1997). RNA polymerase II transcription initiation: a structural view. Proc. Natl. Acad. Sci. U.S.A. 94 (1): 15–22. PMID 8990153. doi:10.1073/pnas.94.1.15.
- Reese JC (April 2003). Basal transcription factors. Current opinion in genetics & development 13 (2): 114–8. PMID 12672487. doi:10.1016/S0959-437X(03)00013-3.
- Shilatifard A, Conaway RC, Conaway JW (2003). The RNA polymerase II elongation complex. Annual review of biochemistry 72: 693–715. PMID 12676794. doi:10.1146/annurev.biochem.72.121801.161551.
- Thomas MC, Chiang CM (2006). The general transcription machinery and general cofactors. Critical reviews in biochemistry and molecular biology 41 (3): 105–78. PMID 16858867.
- Chris Nelson, Susan Goto, Karen Lund, Wesley Hung and Ivan Sadowski (January 2003). Srb10/Cdk8 regulates yeast filamentous growth by phosphorylating the transcription factor Ste12. Nature 421: 187–190. PMID 12520306. doi:10.1038/nature01243.
- Coen, Henrico S.; Elliot M. Meyerowitz (1991). The war of the whorls: Genetic interactions controlling flower development. Nature 353: 31–37. doi:10.1038/353031a0.
- Lemons D, McGinnis W (September 2006). Genomic evolution of Hox gene clusters. Science (New York, N.Y.) 313 (5795): 1918–22. PMID 17008523. doi:10.1126/science.1132040.
- Moens CB, Selleri L (March 2006). Hox cofactors in vertebrate development. Developmental biology 291 (2): 193–206. PMID 16515781. doi:10.1016/j.ydbio.2005.10.032.
- Ottolenghi C, Uda M, Crisponi L, Omari S, Cao A, Forabosco A, Schlessinger D (January 2007). Determination and stability of sex. BioEssays : news and reviews in molecular, cellular and developmental biology 29 (1): 15–25. PMID 17187356. doi:10.1002/bies.20515.
- Toyn JH, Johnson AL, Donovan JD, Toone WM, Johnston LH. (January 1997). The Swi5 transcription factor of Saccharomyces cerevisiae has a role in exit from mitosis through induction of the cdk-inhibitor Sic1 in telophase.. Genetics 145 (1): 85–96. PMID 9017392.
- Dirk Inzé. Cell Cycle Control and Plant Development. Wiley InterScience. ISBN 9780470988923. doi:10.1002/9780470988923.
- Ohta M, Ohme-Takagi M, Shinshi H. (2000). Three ethylene-responsive transcription factors in tobacco with distinct transactivation functions.. Plant J. 22 (1): 29–3. PMID 10792818.
- Nair M, Bilanchone V, Ortt K, Sinha S, Dai X. (Feb 2007). Ovol1 represses its own transcription by competing with transcription activator c-Myb and by recruiting histone deacetylase activity. Nucleic Acids Research 35 (5): 1687–97. PMID 17311813.
- Domian IJ, Reisenauer A, Shapiro L. (1999). Feedback control of a master bacterial cell-cycle regulator.. Proc Natl Acad Sci U S A. 96 (12): 6648–53. PMID 10359766.
- Palatnik JF, Allen E, Wu X, Schommer C, Schwab R, Carrington JC, Weigel D (September 2003). Control of leaf morphogenesis by microRNAs.. Nature 425: 257–263. PMID 12931144. doi:10.1038/nature01958.
- Whiteside ST, Goodbourn S (April 1993). Signal transduction and nuclear targeting: regulation of transcription factor activity by subcellular localisation. Journal of cell science. 104 ( Pt 4): 949–55. PMID 8314906.
- Bohmann D (November 1990). Transcription factor phosphorylation: a link between signal transduction and the regulation of gene expression. Cancer cells (Cold Spring Harbor, N.Y. : 1989) 2 (11): 337–44. PMID 2149275.
- Weigel NL, Moore NL (2007). Steroid Receptor Phosphorylation: A Key Modulator of Multiple Receptor Functions. PMID 17536004. doi:10.1210/me.2007-0101.
- Wärnmark A, Treuter E, Wright AP, Gustafsson J-Å (2003). Activation functions 1 and 2 of nuclear receptors: molecular strategies for transcriptional activation. Mol. Endocrinol. 17 (10): 1901–9. PMID 12893880. doi:10.1210/me.2002-0384.
- Littlewood TD, Evan GI (1995). Transcription factors 2: helix-loop-helix. Protein profile 2 (6): 621–702. PMID 7553065.
- Vinson C, Myakishev M, Acharya A, Mir AA, Moll JR, Bonovich M (September 2002). Classification of human B-ZIP proteins based on dimerization properties. Molecular and cellular biology 22 (18): 6321–35. PMC 135624. PMID 12192032. doi:10.1128/MCB.22.18.6321-6335.2002.
- Wintjens R, Rooman M (September 1996). Structural classification of HTH DNA-binding domains and protein-DNA interaction modes. Journal of molecular biology 262 (2): 294–313. PMID 8831795. doi:10.1006/jmbi.1996.0514.
- Gehring WJ, Affolter M, Bürglin T (1994). Homeodomain proteins. Annual review of biochemistry 63: 487–526. PMID 7979246. doi:10.1146/annurev.bi.63.070194.002415.
- Dahl E, Koseki H, Balling R (September 1997). Pax genes and organogenesis. BioEssays : news and reviews in molecular, cellular and developmental biology 19 (9): 755–65. PMID 9297966. doi:10.1002/bies.950190905.
- Laity JH, Lee BM, Wright PE (February 2001). Zinc finger proteins: new insights into structural and functional diversity. Current opinion in structural biology 11 (1): 39–46. PMID 11179890. doi:10.1016/S0959-440X(00)00167-6.
- Wolfe SA, Nekludova L, Pabo CO (2000). DNA recognition by Cys2His2 zinc finger proteins. Annual review of biophysics and biomolecular structure 29: 183–212. PMID 10940247. doi:10.1146/annurev.biophys.29.1.183.
- http://www.cbrc.jp/research/db/TFSEARCH.html
- Public http://www.gene-regulation.com/pub/programs.html
- Orphanides G, Lagrange T, Reinberg D (1996). The general transcription factors of RNA polymerase II. Genes Dev. 10 (21): 2657–83. PMID 8946909. doi:10.1101/gad.10.21.2657.
- Stegmaier P, Kel AE, Wingender E (2004). Systematic DNA-binding domain classification of transcription factors. Genome informatics. International Conference on Genome Informatics 15 (2): 276–86. PMID 15706513. Архів оригіналу за 19 червня 2013. Процитовано 23 жовтня 2008.
- Matys V, Kel-Margoulis OV, Fricke E, Liebich I, Land S, Barre-Dirrie A, Reuter I, Chekmenev D, Krull M, Hornischer K, Voss N, Stegmaier P, Lewicki-Potapov B, Saxel H, Kel AE, Wingender E (2006). TRANSFAC and its module TRANSCompel: transcriptional gene regulation in eukaryotes. Nucleic Acids Res. 34 (Database issue): D108–10. PMID 16381825. doi:10.1093/nar/gkj143.
- TRANSFAC® database. Архів оригіналу за 21 березня 2012. Процитовано 5 серпня 2007.
- Ben-Gal I, Shani A, Gohr A, Grau J, Arviv S, Shmilovici A, Posch S, Grosse I (2005). Identification of Transcription Factor Binding Sites with Variable-order Bayesian Networks. Bioinformatics 21 (11): 2657–2666. doi:10.1093/bioinformatics/bti410. Архів оригіналу за 4 серпня 2009. Процитовано 24 жовтня 2008.
- Yoshida T, Nishimura T, Aida M, Pichierri F, Gromiha MM, Sarai A. (2001-2002). Evaluation of free energy landscape for base-amino acid interactions using ab initio force field and extensive sampling. Biopolymers 61 (1): 84–95. PMID 11891631.
- Kono H, Sarai A. (1999). Structure-based prediction of DNA target sites by regulatory proteins.. Proteins 35 (1): 114–31. PMID 11891631.
- Transcription Factors Bind Thousands of Active and Inactive Regions in the Drosophila Blastoderm. Архів оригіналу за 25 червня 2013.
- Deng, Q.-L., Ishii, S. and Sarai, A. (1996). Binding-Site Analysis of c-Myb: Screening of Potential Binding Sites by the Mutational Matrix Derived from Systematic Binding Affinity Measurements. Nucleic Acids Res 24: 766–774. PMID 8604322.
- Czechowski T, Bari RP, Stitt M, Scheible WR, Udvardi MK. (2004). Real-time RT-PCR profiling of over 1400 Arabidopsis transcription factors: unprecedented sensitivity reveals novel root- and shoot-specific genes.. Plant J 38 (2): 366–79. PMID 15078338.
- Tabrizifard S, Olaru A, Plotkin J, Fallahi-Sichani M, Livak F, Petrie HT. (2004). Analysis of transcription factor expression during discrete stages of postnatal thymocyte differentiation.. J Immunol 173 (2): 1094–102. PMID 15240698.
- Becker M, Baumann C, John S, Walker DA, Vigneron M, McNally JG, Hager GL. (2002). Dynamic behavior of transcription factors on a natural promoter in living cells... EMBO Rep 3 (12): 1188–94. PMID 12446572.
- Trappe R, Laccone F, Cobilanschi J, et al (May 2001). MECP2 mutations in sporadic cases of Rett syndrome are almost exclusively of paternal origin. American journal of human genetics 68 (5): 1093–101. PMC 1226090. PMID 11309679. doi:10.1086/320109.
- Vaxillaire M, Boccio V, Philippi A, Vigouroux C, Terwilliger J, Passa P, Beckmann JS, Velho G, Lathrop GM, Froguel P (April 1995). A gene for maturity onset diabetes of the young (MODY) maps to chromosome 12q. Nat. Genet. 9 (4): 418–23. PMID 7795649. doi:10.1038/ng0495-418.
- Entrez Gene: PDX1 pancreatic and duodenal homeobox 1.
- Vargha-Khadem F, Gadian DG, Copp A, Mishkin M (2005). FOXP2 and the neuroanatomy of speech and language. Nature Reviews Neuroscience 6: 131–137. PMID 15685218. doi:10.1038/nrn1605.
- National Center for Biotechnology Information. The p53 tumor suppressor protein. Genes and Disease. United States National Institutes of Health. Процитовано 28 травня 2008.
- Gearhart J, Pashos EE, Prasad MK, Pluripotency Redeux — advances in stem-cell research, N Engl J Med 357(15):1469 Free full text
- Wang H, Nussbaum-Wagler T, Li B, Zhao Q, Vigouroux Y, Faller M, Bomblies K, Lukens L, Doebley JF. (2005). The origin of the naked grains of maize.. Nature 436: 714–9. PMID 16079849.
- FRARY, A., T. C. NESBITT, S. GRANDILLO, E. KNAAP, B. CONG et al. (2000). Tfw2.2: A Quantitative Trait Locus Key to the Evolution of Tomato Fruit Size. Science 289: 85 – 88. doi:10.1126/science.289.5476.85.
- SIMONS, K. J., J. P. FELLERS, H. N. TRICK, Z. ZHANG, Y. S. TAI et al. (2006). Molecular Characterization of the Major Wheat Domestication Gene Q. Genetics 172: 547–555. doi:10.1534/genetics.105.044727.