Дикінсонія
Dickinsonia (укр. Дикінсонія) — одна з найхарактерніших викопних тварин едіакарської (вендської) біоти. Як правило, являє собою двосторонньо-симетричне рифлене овальне тіло. Родинні зв'язки організму в даний час невідомі. Більшість дослідників відносять дикінсонію до тварин. Відкриття молекул холестеролу у викопних дикінсоній у 2018 році підтвердило гіпотезу, що вони були найдавнішими серед нині відомих тварин[1][2].
? † ДикінсоніяЧас існування: 575–545 млн р. т. | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
| Dickinsonia costata Sprigg, 1947 | ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||

Дикінсонія і її рідня (так звані проартикуляти) — одні з найбільш дивних і загадкових представників едіакарської біоти, населяла моря 571—541 млн років тому, незадовго до кембрійського вибуху. Проартикуляти так сильно відрізняються від всіх сучасних організмів, що визначити їх місце в системі живої природи довго не вдавалося: трактування варіювали від колоніальних бактерій і гігантських протистів до наземних лишайників і вищих тварин (білатерій). Палеонтологи з Росії, Австралії та Німеччині внесли ясність у це заплутане питання, вивчивши «молекулярні копалини» — залишки еукаріотичних мембранних стеролів, що збереглися у відбитках дикінсоній з місцезнаходження Лямца (Онезький берег Білого моря). Отримані результати дозволяють впевнено віднести дикінсонію (а значить, і її рідню — інших проартикулят) до царства тварин.[3]
Видова різноманітність
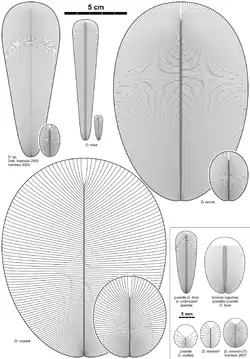
Всього описано 9 видів дикінсоній:
- D. costata Sprigg, 1947
- D. minima Sprigg, 1949
- D. spriggi Harrington et Moore, 1955
- D. elongata Glaessner et Wade, 1966
- D. tenuis Glaessner et Wade, 1966
- D. lissa Wade, 1972
- D. brachina Wade, 1972
- D. menneri Keller 1976, повторно описані Іванцовим в 2007 році[4] (= Vendomia menneri Keller 1976[5])
- D. rex Jenkins, 1992
Надійно виділені тільки 4 або 5 з нижчеперелічених видів:
- D. costata. Синоніми: D. minima, D. spriggi, і D. elongata (голотип). На відміну від інших видів має меншу кількість більш широких сегментів (ізомерів).
- D. tenuis. Синонім: D. brachina. Дуже нагадує D. costata, але має більш численні і вузькі сегменти, овальні обриси тіла.
- D. lissa — подовжений (до 15 см), що нагадує стрічку з безліччю тонких ізомерів, голова і прилеглі до неї ізомери дуже короткі. Скам'янілості мають характерний осьової виступ, що складається з двох смуг, які простягаються від області голови до заднього кінця тіла.
- D. menneri — невеликий організм близько 8 мм у довжину, що нагадує незрілу особину D. costata невеликим числом ізомерів і добре окресленою головою, сформованою сегментованими або злитими ізомерами. Відрізняється від D. costata видовженою формою.
- D. rex — цей вид виділений з D. elongata. Представлений декількома дуже великими екземплярами (до 1 метра і більше в довжину) і не має точного визначення. Основним аргументом для виділення є великі розміри, хоча може бути великим примірником D. costata або / і D. tenuis.
Морфологія
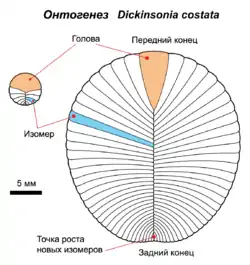
Особи від 4 мм до 1,4 м в довжину[6] овальної форми. Складаються з великої кількості стрічкових сегментів, що виходять з центрального гребеня або жолобка. Напівсегменти (ізомери) обох сторін зрушені відносно один одного на половину ширини, утворюючи симетрію ковзного відображення.
Найбільші екземпляри виявлені в едіакарських відкладеннях в Австралії. На Зимовому березі в Архангельській області знайдено скам'янілості довжиною до 50 см[7].
Сегменти дикінсонії описуються як «пневматики», заповнені рідиною під тиском вище, ніж тиск навколишнього середовища[8]. На деяких примірниках помічені сліди, що інтерпретуються як поздовжні м'язові волокна[9] і медіальна кишка, але ця інтерпретація не загальноприйнята.
Вважають, що нутрощі дикінсонії містять веретеноподібні волокна товщиною 0,5-1 мм[6].
Знахідки скам'янілостей
Dickinsonia вперше описана Реджинальдо Спріггом (Reg Sprigg), першовідкривачем едіакарської біоти в Австралії[10]. Названа ним на честь Бена Дікінсона (Ben Dickinson), директора гірничорудної промисловості Південної Австралії і глави урядового департаменту, де працював Спрігг.
Dickinsonia відома за відбитками в кварцових пісковиках пізнього едіакарського періоду[6] в Едіакарії та інших місцях хребта Фліндерс в Південній Австралії, а також на Поділлі в Україні, на узбережжі Білого моря і на центральному Уралі в Росії. Вік скам'янілостей оцінюється в 560—555 млн років[11].
Дикінсонія є «стійкою» скам'янілістю, на відміну від інших едіакарських організмів вона зберігається як увігнутий відбиток в вищерозміщених нашаруваннях. Якщо зберігся і верхній, і нижній відбиток, вони зазвичай відстоять по вертикалі на відстань близько 3 мм з найсильнішим рифленням на верхньому відбитку, що дозволяє припустити, що рельєф був тільки на спинній поверхні організму.
Сліди
Знайдено дугоподібні сліди дикінсонії, віднесені до іхнотаксона[12], проте їх інтерпретація не безперечна. Це могли бути слизові виділення організму на підстилаючих мікробних матах[6], можливо, з метою перетравлювання[6][13]. Деякі дослідники інтерпретують ці скам'янілості як сліди перекочування по дну якогось предмета або організму під дією потока води[6] або відбитки, залишені групою лишайників або грибів (на зразок «відьомських кіл»)[6]. Проте в деяких випадках сліди перекривають один одного. Спостерігаються доріжки, сформовані, ймовірно, кишковими виділеннями, що підтверджують версію, що сліди залишені в процесі харчування. До того ж порушення структури осадових порід, викликані падінням або перекочування по дну, не виявлені[14].
Взаємодія скам'янілостей
Не виявлено жодного випадку, коли б скам'янілості окремих організмів перекривалися. Навколо кожного організму спостерігається своєрідна «смуга відчуження». Організми, що знаходяться в безпосередній близькості один від одного, часто деформовані, щоб не порушити її[6].
Внутрішня анатомія

Деякі скам'янілості дикінсонії зберегли елементи внутрішньої анатомії. Припускається, що вони являють собою тракт, що забезпечує перетравлювання їжі і розподіл поживних речовин по організму[15]. На одному з родовищ Зимового берега в Архангельській області знайдено понад 20 відбитків дикінсонії зі слідами шлунково-кишкового тракту[7].
Тафономія
Дикінсонії, як і інші мешканці едіакарської біоти, не мали мінералізованого скелета (раковини або панцира), їх тіло складалося тільки з м'яких тканин, подібно до сучасних медуз або слимаків. З цієї причини вендські скам'янілості представлені тільки відбитками на мілководних піщано-глинистих відкладах. Виникнення відбитків пов'язують з лавиноподібним накопиченням опадів під час шторму або підводного зсуву. Коли ділянка дна накривається новим осадовим шаром, на підошві останнього утворюється відбиток похованих ним живих організмів. Розрізняють такі типи відбитків[7]:
- Негативні (вдавлені в шар), що представляють відбитки верхньої частини тварин, накритих і здавлених осадовим шаром;
- Позитивні (підносяться над шаром), що являють собою вм'ятини на ґрунті, заповнені вищерозташованими осадовими породами. Зазвичай це сліди тварин або відбитки нижньої частини їхніх тіл.
Завдяки тому, що різні частини тіла мали різну щільність і розкладалися з різною швидкістю, в рельєфі відбитків виявляється не тільки топографія зовнішньої поверхні організму, але і елементи його внутрішньої будови. Стійкіші тканини і органи утворювали виступи, менш стійкі — западини[7].
Велика кількість відбитків м'якотілих тварин є унікальною особливістю едіакарського періоду. В едіакарській біоті відсутні трупоїди, подрібнювачі мертвих тіл і біотурбатори (риючі тварини), що руйнували відбитки на м'якому ґрунті. Мертва органіка розкладалася тільки завдяки життєдіяльності мікробів[7].
У тих зразках, де дикінсонія складена або зігнута, вона демонструє відсутність руйнувань, характерних для тендітних структур[6].
Товщина знайдених зразків мало залежить від лінійних розмірів, що характерно швидше для лишайників, листяних рослин або грибів, ніж для м'якотілих тварин[6]. Вважають, що дикінсонії були жорсткішими, ніж черв'яки або медузи завдяки гідростатичному скелету[16].
У виявлених скупченнях організмів присутні особини різних розмірів, що свідчить про їх поховання в природних умовах. У скупченнях, утворених наносами водяних потоків переважали б особини менших розмірів[6]. Виявлення деяких екземплярів на вершинах осадових структур свідчить про їх надійне прикріплення до дна в момент поховання[6].
Дикінсонії виявляються в осадових шарах товщиною від 8 мм. Це свідчить про те, що їхня товщина з урахуванням здавлювання не перевищувала 1 см.
Екологія
У дикінсоній спостерігається необмежений ізометричний ріст — збільшення геометричних розмірів із збереженням пропорцій до тих пір, поки організм не буде похований осадовими породами або убитий яким-небудь іншим чином[6]. Велику частину життя вони, ймовірно, проводили в нерухомості, прикріпленими до донних опадів, однак могли пересуватися з місця на місце. Тип прикріплення до субстрату, можливо, схожий зі зрощенням вустриць, ризоїдами лишайників або подобою підземної грибниці[6]. Збереження організмів свідчить, що несучими елементами скам'янілостей були скоріше біополімери типу кератину, ніж тендітні мінерали типу кальциту чи посмертна піритізація тіла.
Родова подібність
Родинні відносини дикінсонії з сучасними типами тварин неясні. Різні дослідники відносять її до медуз, коралів, актиній, ксенофіофор, поліхет багатощетинкових або війчастих черв'яків, грибів, лишайників[6][17] і навіть близьких попередників хордових[18].
Реталлек зробив спробу простежити родинні стосунки дикінсоній, використовуючи схожість процесу розкладання решток і припустив, що організми едіакарської біоти були споріднені лишайникам[16], проте його погляди зазнали різкої критики[19]. Тим не менш, режим розкладання едіакарських організмів дійсно більш схожий з процесом розкладання листяних рослин, грибів і лишайником, ніж м'якотілих тварин, які в процесі розпаду стискаються і деформуються[6].
Довгі суперечки про родинні зв'язки едіакарських організмів привели до погляду (який був вперше висловлений Раннегаром в 1958 році), що вони відносяться до особливого царства живої природи, «вендобіонтів» або «вендозоїв», які вимерли на межі кембрійського періоду і не залишили нащадків серед сучасних типів організмів[20]. Інші вендобіонти (Йоргія, Marywadea) мають багато схожих рис з дикінсоніями і, мабуть, перебували з ними у родинних стосунках. Основні відмінності дикінсоній від тварин сучасних типів наступні[21]:
- Ізометричний ріст, тобто зростання зі збереженням пропорцій організму, тоді як у тварин сучасних типів пропорції тіла різняться у дитинчат і дорослих особин[21];
- Своєрідний тип симетрії, схожий з білатеральною симетрією сучасних організмів, однак із зсувом правих і лівих напівсегментів відносно один одного. М. О. Федонкін позначив цей тип симетрії запозиченим з математики терміном «симетрія ковзного відбиття». У сучасних тварин ковзна симетрія зустрічається вкрай рідко (один із прикладів — розташування м'язових елементів — міомерів і міосепт — у ланцетника)[21].
Відзначено, що симетрія ковзного відображення характерна для листкоподібних органів папороті, у зв'язку з чим деякі дослідники відносять дикінсонію і схожі на неї організми (наприклад, Чарнію) в таксон Rangeomorph (рангеоморфів)[22].
Загальновизнано, що дикінсонії могли пересуватися, про що свідчать їхні сліди на бактеріальних матах, залишені в процесі харчування[23]. Однак немає безперечних свідчень наявності у дикінсоній рота, кишечника і анального отвору. Ймовірно, дикінсонії харчувалися за допомогою поглинання поживних речовин нижньою поверхнею тіла і, таким чином, вони займають проміжне положення між губками та еуметазоями і, можливо, споріднені пластинчастим[23][24].
Примітки
- Bobrovskiy, Ilya; Hope, Janet M.; Ivantsov, Andrey; Nettersheim, Benjamin J.; Hallmann, Christian; Brocks, Jochen J. (20 вересня 2018). Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals. Science 361 (6408): 1246–1249. doi:10.1126/science.aat7228.
- Учені знайшли залишки найстарішої тварини. Їй 558 млн років. Українська правда. 24 вересня 2018. Процитовано 24 вересня 2018.
- Ilya Bobrovskiy, Janet M. Hope, Andrey Ivantsov, Benjamin J. Nettersheim, Christian Hallmann, Jochen J. Brocks. Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals // Science. — 2018. — V. 361. — P. 1246—1249.
- Ivantsov, A. Yu. Small Vendian transversely Articulated fossils // Paleontological Journal. – 2007. – Vol. 41, Issue 2. – P. 113-122
- Keller B. M., Fedonkin B. M. New Records of Fossils in the Valdaian Group of the Precambrian on the Syuz'ma River // Izv. Akad. Nauk SSSR, Ser. Geol. – 1976. – Vol. 3. – P. 38-44
- Retallack G.J. Growth, decay and burial compaction of Dickinsonia, an iconic Ediacaran fossil // Alcheringa: an Australasian Journal of Palaeontology. - 2007. – Vol. 31, Issue = 3. – P. 215-240
- Малаховская Я. Е., Иванцов А. Ю. Вендские жители Земли. - Химия и жизнь. – 2004. - № 7.
- Seilacher 1989
- Dzik 2000??
- Sprigg Reg C. Early Cambrian (?) Jellyfishes from the Flinders Ranges, South Australia // Trans. Roy. Soc. S. Aust. – 1947. – Vol. 71. – P. 212-24
- Grazhdankin Dima. Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution // Palæobiology. — 2004. — Vol. 30, Issue 2. — P. 203—221
- Ivantsov A.Y., Malakhovskaya Y.E. Giant Traces of Vendian Animals // Doklady Earth Sciences. — 2002. — Vol. 385, Issue 6. — P. 618—622
- Jensen Sören, Droser Mary L.; Gehling James G. Trace fossil preservation and the early evolution of animals // Palaeogeography, Palaeoclimatology, Palaeoecology. – 2005. – Vol. 220, Issue 1-2. – P. 19-29
- Ivantsov A. Yu. Feeding traces of Proarticulata - the Vendian metazoan // Paleontological Journal. – 2011. – Vol. 45, Issue 3. – P. 237-248
- Ivantsov A. Yu. New Proarticulata from the Vendian of the Arkhangel'sk Region // Paleontological Journal. – 2004. – Vol. 38, Issue 3. – Р. 247-253
- Retallack G.J. Were the Ediacaran fossils lichens? // Paleobiology. – 1994. – Vol. 17. – P. 523-544
- Retallack Gregory J. Decay and Destruction of Dickinsonia // Geological Society of America. Abstracts with Programs. – 2004. - Vol. 36, No. 5. - Р. 521
- Dzik Jerzy. The Origin of the Mineral Skeleton in Chordates // Evolutionary Biology / Max Knobler Hecht, Ross J. MacIntyre and Michael T. Clegg, eds. – 2000. - Vol. 31. - P. 105-46.
- Waggoner B., Collins A.G. Reductio Ad Absurdum': Testing The Evolutionary Relationships Of Ediacaran And Paleozoic Problematic Fossils Using Molecular Divergence Dates. - Journal of Paleontology. – 2004. – Vol. 78, Issue = 1. – P. 51-61
- Runnegar Bruce. Oxygen requirements, biology and phylogenetic significance of the late Precambrian worm Dickinsonia, and the evolution of the burrowing habit // Alcheringa: An Australasian Journal of Palaeontology. – 1982. – Vol. 6. – P. 223-239
- Такономическое положение вендобионтов (представителей эдиакарской фауны) - на сайте medbiol.ru.
- Brasier M., Antcliffe J. Decoding the Ediacaran Enigma // Science. – 2004. – Vol. 305, Issue 5687. – P. 1115-1117
- Sperling Erik, Vinther Jakob, Pisani Davide, Peterson Kevin. A placozoan affinity for Dickinsonia and the evolution of Late Precambrian metazoan feeding modes. // Cusack M., Owen A., Clark N. Programme with Abstracts. 52. Palaeontological Association Annual Meeting. - Glasgow, UK. – 2008. - P. 81.
- Sperling Erik et al. A Placozoan Affinity for Dickinsonia and the Evolution of Late Precambrian Metazoan Feeding Modes // Geological Society of America Abstracts with Programs. - 2008. – Vol. 40, Issue 6. – P. 508