Водорості
Водорості (лат. Algae) — гетерогенна група еукаріотів, що охоплює кілька різних груп відносно простих за структурою живих організмів, які отримують необхідну для життєдіяльності енергію шляхом фотосинтезу, мешкають переважно у водному середовищі чи пристосувались до життя у ґрунті та інших наземних місцях зростання. Водорості традиційно розглядалися як нижчі рослини; деякі з них (зелені водорості) близько споріднені з вищими рослинами. Інші представники належать до різних груп найпростіших. Деякі групи подібні до тварин (наприклад, Apicomplexa). Різні групи водоростей виникли у різний час від різних предкових форм (поліфілетично) та розвивалися окремо, але в результаті конвергентної еволюції набули багато схожих рис.
? Водорості | ||||||
|---|---|---|---|---|---|---|
 Водорості різних видів | ||||||
| Біологічна класифікація | ||||||
| ||||||
Включені групи | ||||||
| ||||||
Посилання | ||||||
| ||||||
| Ботаніка |
|---|
 Різноманіття рослинного світу |
|
Знамениті ботаніки |
|
Історія
|
|
|
Наука про водорості називається альгологією.
Загальні відомості

Водорості — поліфілетична група організмів, які об'єднані за такими ознаками: наявність фотосинтетичних пігментів, що визначають їх різноманітне забарвлення, та здатність до оксигенного фотосинтезу. Окрім того, до водоростей включають деякі безколірні організми, що вторинно втратили фотосинтетичні пігменти (теорія ендосимбіозу) та, відповідно, здатність до фототрофного живлення, проте мають з водоростями близьку генетичну спорідненість. Деякі водорості здатні до гетеротрофії, як осмотрофно (поверхнею клітини), так і шляхом активного захоплення їжі (деякі евгленові, динофітові). Розміри водоростей коливаються від мікрона (більшість одноклітинних водоростей) до 40 м (Бурі водорості). Талом — вегетативне тіло водоростей, може бути як одно-, так і багатоклітинним. Серед багатоклітинних є й одноклітинні[неоднозначно – уточнити] (наприклад, спорофіт бурих водоростей). Серед одноклітинних є колоніальні форми — певна парна кількість клітин, що з'єднані між собою через плазмодесми або оточені загальним слизом.
Серед водоростей на сьогодні[коли?] описано понад 60 тис. видів, разом з тим очікуване різноманіття за оцінками різних авторів[яких?] лежить у межах 500 тис. — 10 млн видів. Об'єднують водорості у різну кількість видів.[джерело?]
Тривалий час до водоростей традиційно включали також ціанобактерій («синьозелені водорості»), які належать до прокаріот, але мають досить великі клітини та морфологічну подібність до водоростей.
Деякі ботаніки, віддаючи данину традиції, схильні й далі вважати ціанобактерії водоростями. Альгологи описали понад 1000 видів синьозелених, проте станом на 2019 рік бактеріологічними методами підтверджено існування не більше 400 штамів[1].
Цитологія
Цитологічні ознаки водоростей є цілком типовими для інших еукаріот (наявність ядра, комплексу Ґольджі, мітохондрій). Разом з тим вони мають специфічні ознаки, що властиві лише рослинним (наявність клітинної оболонки, хлоропластів, вакуолей та ін.) чи лише тваринним організмам (руховий апарат, центріолі). Синьозелені водорості за будовою нагадують бактерії, разом з якими їх відносять до без'ядерних (прокаріотичних) організмів.
До цитологічних ознак, що є систематичними ознаками на рівні відділів, належать такі:
- ознаки, що відображують спорідненість таксонів за клітиною-господарем (покриви, ядерний апарат);
- система мікротрубочкових органел (джгутиковий апарат)
- мітохондріальний апарат.
Типи морфологічної структури тіла водоростей
Найперше різноманітність водоростей простежується у зовнішньому вигляді. Вони можуть мати зовсім просту будову і зовсім незначні розміри, або навпаки, величезні розміри та тканинну будову. Так звані плани будови водоростей, що мають важливу діагностичну ознаку, називають типами морфологічної структури. Різні автори виділяють різну кількість таких типів:
- монадний
- гемімонадний
- кокоїдний
- нитчастий (трихальний)
- різнонитчатий (гетеротрихальний)
- тканинний
- сифональний
- сифонокладальний
Розмноження та цикли розвитку
Розмноження у водоростей відбувається двома шляхами: статевим та нестатевим. Відповідно при нестатевому розмноженні дочірні особини успадковують такий геном, що є ідентичним до батьківського геному, за винятком мутаційних процесів. При статевому розмноженні дочірні геноми відрізняються від батьківських, через те, що утворенню нової генерації передує процес кросинговеру.
Нестатеве розмноження водоростей умовно можна поділити на два типи:
Споруляція
Нестатеве розмноження шляхом споруляцiï здiйснюється спеціалізованими клітинами - спорами, які утворюються в спорангіях. Спорангії у водоростей одноклітиннi, кiлькiсть спор в одному спорангії може коливатися від однієï (наприклад, у зеленої нитчастої водорості Oedogonium) до кількох десятків або навіть сотень (у багатьох бурих водоростей).
У відповідності зі здатністю до активного руху спори поділяють на рухливі із джгутиками - зооспори, нерухомi з клітинними покривами гемiзооспори, апланоспори та автоспори, та малорухливі амебоїдні, що позбавлені джгутиків - моноспори, бiспори та тетраспори.
Зооспори завжди мають джгутики і часто - вічко та пульсуючi вакуолі. Зооспори можуть бути голими або вкритими клітинними покривами.
Гемiзооспори джгутикiв не мають, проте зберігають ознаки монадної організації - стигму, пульсуючі вакуолі, базальнi тiла джгутиків. Зазвичай гемiзооспори спостерігаються в амфібіальних водоростей, які у водному середовищі утворюють зооспори, проте у позаводному припиняють процес формування джгутиків на стадії їх базальних тіл.
Апланоспори джгутикiв також не мають, їх початкові етапи розвитку нагадують ранні стадії процесу утворення зооспор, зокрема, в протопластах майбутніх апланоспор утворюються базальні тіла. Проте на кінцевих етапах розвитку, незалежно від наявності у середовищі води, базальнi тiла або зникають, або мігрують вглиб клітини і перетворюються на центріолі. Інколи в апланоспор можна спостерігати пульсуючі вакуолі, проте стигми у них не розвиваються. Зазвичай зрілі апланоспори не повторюють морфології материнських клітин.
Автоспори у своєму розвитку не проходять навіть початкових стадій зооспороутворення. В автоспор відсутні джгутики, їх базальні тiла, стигми, пульсуючі вакуолі. Морфологічно автоспори подібні до материнських клітин і відрізняються від них лише меншими розмірами.
Апланоспори та автоспори, крім функції розмноження, часто здатні виконувати й функцію перенесення несприятливих умов існування. Амебоїдні безджгутикові спори - моноспори, бiспори, тетра спори - відомі у червоних та деяких бурих водоростей. Клітина такої спори являє собою голий протопласт, одягнений лише плазмалемою і здатний до повільного амебоїдного руху. Утворенню цих спор у переважній більшості випадків передує мейоз, через що моно-, бі- та тетраспори мають гаплоїдний набір хромосом. Назви цих спор вiдображають те, в якій кількості утворюються спори у спорангії - по одній (моноспори), по дві (біспори) чи по чотири (тетраспори).
За типом поділу ядра в клітині-спорангії (мітотичний чи мейотич ний) спори також часто поділяють на мітоспори та мейоспори.
Вегетативне розмноження
В одноклітинних водоростей вегетативне розмноження бувається переважно поділом клітини на двоє. У багатоклітинних та колоніальних представників вегетативне розмноження частіше пов'язане з фрагментацією таломів та колоній. Фрагментацію можуть спричинювати механічні чинники (наприклад, вітро-хвильові удари, течія) або фрагментація може відбуватися внаслідок відмирання частин таломів чи колоній. Наприклад, відмирання окремих інтеркалярних клітин у багатоклітинних трихомах синьо-зелених водоростей часто обумовлює розпад трихому на репродуктивні фрагменти - гормогонії. У небагатьох водоростей вегетативне розмноження може здійснюватися за допомогою спеціалізованих ділянок тіла - вивідкових бруньок (бура водорість Sphacellaria) або вивідкових бульбочок (зелені водорості з порядку Charales). Особливим варіантом вегетативного розмноження є розмноження за допомогоюакінет. Останні являють собою клітини, що утворюються із звичайних вегетативних клітин при погіршенні умов існування. Акінети мають потовщену клітинну оболонку, містять багато запасних поживних речовин і, на відміну від нормальних вегетативних клітин, за несприятливих умов здатні переходити у стан анабіозу або криптобіозу, і завдяки цьому переживати несприятливі періоди. При покращенні умов вегетації акінети проростають у нові вегетативні особини.
Статеве розмноження
Статеве розмноження відбувається внаслідок статевого процесу, який створює умови для кросинговеру, що, у свою чергу, забезпечує певний рівень генетичноï гетерогенностi популяцій на основі рекомбінантної мінливості. Статеві процеси у водоростей поділяють на дві основні групи - соматогамнi та гаметогамнi.
При соматогамних статевих процесах зливаються дві вегетативні (інакше - соматичні) клітини. Якщо такі клітини представленi монад ними клітинами, позбавленими клітинних покривів, то цей тип сома тогамiї називають хологамією.
Якщо копуляція відбувається між вегетативними клітинами, які мають клітинні покриви, то цей тип соматогамії називають кон'югацiєю. При кон'югації клітини, що мають копулювати, зближуються та вкриваються спільним слизом. Далі кожна клітина утворює копуляцiйний відросток в напрямку до іншоï. Коли копуляцiйнi вiдростки стикаються один з одним, оболонка між ними розчиняється й утворюється кон'югацiйний канал, по якому ядро з однієї клітини переходить в іншу. Далі ядра зливаються і утворюється зигота.
При гаметогамії копуляція відбувається між спеціалізованими клітинами статевого розмноження - гаметами, які утворюються в статевих органах - гаметангіях. У відповідності з морфологією гамет виділяють три основі типи гаметогамії: iзогамiю, гетерогамію та оогамію.
Ізогамiєю називають такий статевий процес, при якому копулюють дві монадні гамети, які морфологічно виглядають однаковими. Проте фізіологічно та на ультратонкому рівні гамети розрізняються: одна з гамет, що при копуляції приймає ядро іншої гамети, вважається жіночою і позначається знаком «+». Друга гамета, ядро якої при копуляції мігрує у цитоплазму жіночої гамети, вважається чоловічою і позначається знаком «-».При гетерогамії (синонім - анізогамії) копулюють дві монадні гамети, які відрізняються розмірами та ступенем рухливості. Чоловіча гамета дрібна, рухається дуже швидко. Жіноча гамета має бiльшi розміри, нiж чоловіча, і її рухливість обмежена.
Оогамія - це статевий процес, при якому копуляцiя вiдбувається між великою, нерухомою та позбавленою джгутиків жіночою гаметою - яйцеклітиною та дрібною чоловічою гаметою. Чоловічі гамети, що мають джгутики і здатні до активного руху, називають сперматозоїтдами. Якщо ж чоловіча гамета позбавлена джгутиків і є нерухомою, то її називають спермацієм. Гаметангії, в яких утворюються яйцеклітини, називають оогоніями, а сперматозоїди чи спермації - антеридіями. У водоростей оогонії та антеридії, на відміну від гаметангіїв вищих рослин, є одноклітинними (за винятком деяких зелених водоростей з класу харофiцiєвих).
Крiм основних типів гаметогамiï (тобто iзогамії, гетерогамії та оогамії), у водоростей відомо кілька додаткових специфічних типів статевого процесу. Наприклад, у діатомових водоростей дуже поширеним типом статевого процесу є автогамія. При автогамії диплоїдне ядро клітини редукційно ділиться, далi з чотирьох дочірніх гаплоїдних ядер два дегенерують, а два iнших зливаються дають початок ДИПЛОЇДНОМУ Ядру зиготи. Таким чином, при автогамії гамети не утво рюються, а статевий процес відбувається без статевого партнера.
Інший специфічний варіант статевого процесу - атактогамія, яка спостерігається у деяких зелених вольвокальних водоростей (на приклад, у роду Chlorogonium). Тут в гаметангіях представників навіть однієї популяції утворюються монадні гамети рiзних розмірів і копулюють як морфологічно однакові, так і морфологічно відмінні гамети. Причому гамета, що приймає ядро вiд iншої (тобто фізіологічно жiноча гамета), може за розмірами дорівнювати чоловічій, бути більшою від неї або меншою.
У водоростей, що мають iзогамні статеві процеси, розрізняють види гомоталічні та гетероталічні. У гомоталічних видiв як «+», так і «-» гамети утворюються на одному таломі і здатні копулювати між собою, тобто такі види є фізіологічно двостатевими. У гетероталічних видiв «+» та «-» гамети розвиваються на різних таломах, і тому гетероталiчнi представники фізіологічно є роздільностатевими (одностатевими).
Водорості, що мають статеві процеси гетеро- та оогамного типу, поділяють також на однодомні та дводомні. В однодомних водоростей антеридії та оогонії розміщуються на одному таломі, тобто такі види є і фізіологічно, і морфологічно двостатевими. У дводомних видів жiночі та чоловічі статеві органи утворюються на різних таломах, тоб то має місце фізіологічна та морфологічна роздільностатевість (одностатевість).
Після злиття цитоплазм вегетативних клітин чи гамет, що прий мали участь у статевому процесі (т. зв. плазмогамії) зазвичай відбувається злиття ядер - каріогамія - і утворюється диплоїдне ядро зиготи. Проте з цього правила у водоростей відомі й виключення. Наприклад, у багатьох золотистих водоростей після плазмогамії ядра одразу не зливаються, і, як наслідок, утворюється клітина, що містить два генетично відмінні ядра - дикаріон. Така дикарiонтична клітина надалі перетворюється на дикаріонтичну зиготичну цисту, що переходить у стан спокою. Каріогамія і утворення власне зиготи відбувається лише після закінчення періоду спокою.[2]
Походження, філогенія та еволюція
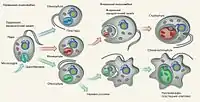
Згідно з сучасною системою органічного світу, водорості є в обох надцарствах органічного світу: Procaryota та Eucaryota. Питання про походження водоростей тривалий час було дискусійним[3][1], власне, як і питання про походження еукаріот. У наш час[коли?] загальновизнаною теорією є синтетична гіпотеза походження еукаріот. Ця теорія об'єднує уявлення щодо автогенетичного походження клітини (шляхом дарвінівської еволюції) та ендосимбіотичну гіпотезу (виникнення евкаріотичної клітини шляхом серії ендосимбіозів між різними організмами). Так, було доведено автогенетичне походження ядра та одномембранних органел та ендосимбіотичне походження пластид та мітохондрій. Виникнення різних груп водоростей відбувалось, очевидно, різними шляхами, про що свідчать результати молекулярно-генетичних, морфологічних та біохімічних досліджень.
Значення у природі
Водорості відіграють важливу роль у синтезі органічної речовини на Землі. У комплексі організмів, які здійснюють колообіг речовин у природі (продуценти — консументи — редуценти), водорості разом з автотрофними бактеріями та вищими рослинами складають ланку продуцентів, за рахунок яких існують усі безхлорофільні організми планети.
| Відділ | Хлорофіли | Каротиноїди | Фікобіліпротеїни | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Синьозелені (разом із прохлорфіолетові) | ||||||||||||
| Глаукоцистофіти | ||||||||||||
| Червоні | ||||||||||||
| Криптофіти | ||||||||||||
| Динофіти | ||||||||||||
| Примнезіофіти | ||||||||||||
| Охрофіти | ||||||||||||
| Евгленові | ||||||||||||
| Хлорарахніофіти | ||||||||||||
| Зелені | ||||||||||||










"" — пігмент відсутній; — пігмент зустрічається рідко або у слідових кількостях; 1 — лише у прохлорофітів; 2 — лише у декотрих прохлорофітів; 3 — лише в Acaryochloris marina; 4 — лише у тих, у яких хлоропласт — ендосимбіотична зелена водорість; 5 — лише у тих, у яких хлоропласт — ендосимбіотична охрофітова водорость; 6 — відсутня в евстигматофіцієвих; 7 — відсутній у трибофіцієвих й евститматофіцієвих; 8 — склад каротиноїдів погано вивчений; 9 — лишу у декотрих празинофіцієвих. [4]
Водорості виступають продуцентами також у наземних місцезростаннях, зокрема найчастіше у таких, що є малопридатними для життя вищих рослин. У таких біотопах вони створюють умови для існування бактерій, грибів та тварин. Наприклад, у водоймах водорості є основним джерелом харчування безхребетних тварин, якими, своєю чергою, харчуються хребетні, в тому числі й риби. Цілком очевидно, що продуктивність зарибленої водойми залежить від продуктивності її водоростевої складової. Беручи участь у процесах колообігу речовин у природі, водорості є активними агентами самоочищення водойм, а також первинних ґрунтотвірних процесів і відновлення родючості ґрунтів.
У геологічному літописі нашої планети водорості також залишили відбиток у вигляді строматолітів, діатомітів та вапняків.
Екологія
Можна виділити такі основні угруповання водоростей:
Планктонні водорості (фітопланктон)

Мешкають у товщі води у завислому стані, для чого зазвичай мають спеціальні морфологічні пристосування. Розповсюджені повсюдно: від калюжі до океану. Відсутні лише у гарячих термальних джерелах та у чистих прильодовикових водах. Не знайдені також у печерних озерах та на великій глибині, де відсутня достатня кількість сонячної енергії для фотосинтезу. Вони є основним, а у деяких випадках — єдиним продуцентом органічної речовини, на основі якого у водоймі існує все живе.
Нейстонні водорості
Мешкають у поверхневій плівці води та не є планктонними організмами. Деякі нейстонні організми знаходяться на поверхневій плівці (епінейстон), інші — під нею (гіпонейстон). У деяких випадках нейстонні організми розвиваються у таких кількостях, що покривають воду суцільною плівкою і мають вирішальне значення у подальшій долі водойми.

Бентосні водорості (фітобентос)
Розвиток водоростей у конкретному місцезростанні зумовлений як абіотичними, так і біотичними факторами. Різні види бентосних водоростей ростуть на різній глибині та у різних гідрохімічних та гідрологічних умовах на дні водойми у вільному або прикріпленому стані. Умовно розрізняють такі екологічні групи бентосних організмів: епіліти (на твердому субстраті) та епіпеліти (на м'якому субстраті), епіфіти (на поверхні рослин), ендоліти (у вапнякових відкладах), а також паразитичні та ендосимбіотичні форми. У континентальних водоймах серед бентосних водоростей переважають діатомові, зелені, синьозелені та жовтозелені водорості, тоді як основними мешканцями океанів та морів є бурі та червоні водорості.
Перифітонні водорості
Мешкають на поверхні різних субстратів, на межі рідкої та твердої фаз. Як субстрати для обростання можуть виступати інші водорості, водні тварини та рослини та їх рештки, а також штучні та природні тверді мінеральні субстрати. Зазвичай можуть мати спеціальні органи для прикріплення, утворення яких зумовлене як морфологічно (різноманітні вирости) та хімічно (слиз). Поряд із молюсками створюють певні труднощі при експлуатації суден та гідротехнічних споруд.

Наземні водорості та лишайники
Мешкають поза водоймами. Протягом майже всього життя оточені повітрям, тому їх ще називають аерофітними. Поселяються на стовбурах дерев, кущів та кущиків, скелях та камінні. Мають пристосування до виживання у наземному середовищі, мешкають у симбіотичних асоціаціях з мікобіотою (лишайники).
Ґрунтові
Мешкають як на поверхні ґрунту, утворюючи іноді розростання у вигляді плівок, так і у глибині ґрунту, де зустрічаються на глибині 2-2,7 м. Найкраще розвиваються біля поверхні на глибині 0,2-1,0 см. Перш за все, це пов'язано з такими необхідними умовами, як вологість, аерація та освітленість. Відіграють велику роль у ґрунтотвірних процесах.
Водорості гарячих вод
Вегетують за температури від +35 °C до +52 °C, в окремих випадках +75 °C, +80 °C та до +84 °C, часто за умов підвищеної концентрації органічних речовин та мінеральних солей.
Водорості солоних водойм

Вегетують як при слабкій, так і підвищеній концентрації солей у воді (іноді до 285–347 г/л). Деякі з них (наприклад, Dunaliella salina) використовують для промислового культивування для отримання каротиноїдів.
Водорості вапнякових відкладів
Мешкають як у водному середовищі, так і поза ним. Можуть руйнувати (шляхом виділення органічних кислот) та створювати (виділяючи кальцій) вапнякові породи.
Співіснування з іншими організмами
Оскільки водорості є широко розповсюдженими організмами, вони вступають у різні форми взаємовідносин з іншими організмами. Окрім взаємодій водоростей із рослинами, грибами (лишайники) та власне водоростями, вони живуть і на тваринах. Наприклад, є симбіонтами, що мешкають у клітинах інших організмів, зокрема, безхребетних. Так, зелена водорість роду Хлорела (Chlorella) поселяється у вакуолях інфузорії Paramecium bursaria. Цікаве екологічне угруповання складають епізоїти, що мешкають на ракоподібних, коловертках та ін. Паразитичні водорості, що мешкають у кишечниках червів, нематод, амфібій, відомі серед євгленофітових та динофітових водоростей.
Застосування

Важливо зазначити їх пряме та непряме значення у медицині, техніці, хімічній промисловості. Умовно можна виділити такі сфери застосування водоростей:
Харчова
Людина здавна використовує водорості у харчовій промисловості, а саме у сфері рибного та сільського господарства. По океанічних та морських узбережжях морські водорості вживають у їжу майже повсюдно, але у Японії вони є національною стравою. У європейських країнах водорості широко використовують як харчову добавку. Найпопулярнішою є так звана морська капуста. Головним чином, це ламінарія та близькі до неї види бурих водоростей. Продукти, що отримують з цих водоростей, у Японії відомі під назвою «комбу». Найчастіше вживають замість звичайної капусти у супах, з рибними та м'ясними стравами. Вживають і у кондитерських виробах.
Окрім морських водоростей, у їжу вживають також прісноводні та наземні водорості, як-от Nostoc pruniforme (носток сливовидний), Nostoc commune (носток звичайний), Nostoc flagelliforme (носток повстяний).
Фармацевтична
З водоростей отримують також агар-агар, агароїди та альгінати. Використовують як джерело каротиноїдів (Dunaliella salina) та харчового йоду (морські водорості).
Технічна
Водорості використовують у космічній техніці, виробництві біопалива та як агенти очистки стічних вод, модельних об'єктів для оцінки стану навколишнього середовища. Є й інша сторона: це обростання гідротехнічних споруд, забивання фільтрів насосних станцій та ін. Розглядаються як перспективні об'єкти культивації для очищення газів та отримання біомаси (отримання целюлози) та олії (отримання біодизелю).
Науково-дослідна
Водорості завдяки особливостям життєвого циклу, клітинної будови, високій продуктивності та зручності культивування (більшості водоростей) є досить зручними модельними об'єктами біологічних досліджень різних напрямів — від вузько спеціалізованих (філогенія водоростей, дослідження цитології та молекулярної біології, дослідження фотосинтезу), так і прикладних (наприклад, для застосування у техніці).
Класифікація
Порядок Chroococcales
- Рід Microcystis
- Рід Merismopedia
- Рід Gleocapsa
Порядок Oscilatoriales
- Рід Oscillatoria
- Рід Arthrospira
Порядок Nostocales
- Рід Gleotrichia
- Рід Aphanizomenon
- Рід Anabena
- Рід Nostoc
Порядок Stigonematales
- Рід Stigonema
Порядок Euglenales
- Рід Euglena
- Рід Trachelomonas
- Рід Phacus
Порядок Peranematales
- Рід Peranema
Порядок Euglenomorphales
- Рід Euglenomorpha
Порядок Chlorarachniales
- Рід Chlorarachnion
Порядок Raphidiales (Vacuolariales)
- Рід Goniostomum
- Рід Vacuolaria
Порядок Ochromonadales
- Рід Ochromonas
- Рід Chromulina
- Рід Dinobryon
Порядок Synurales
- Рід Mallomonas
- Рід Synura
Порядок Eustigmatales
- Рід Eustigmatos
Порядок Vaucheriales
- Рід Vaucheria
Порядок Botrydiales
- Рід Botrydium
- Рід Botrydiopsis
Порядок Mischococcales
- Рід Characiopsis
- Рід Heterococcus
Порядок Tribonematales
- Рід Tribonema
Порядок Laminariales
- Рід Laminaria
- Рід Macrocystis
Порядок Fucales
- Рід Fucus
- Рід Cystoseira
- Рід Sargassum
Порядок Melosirales
- Рід Melosira
Порядок Fragilariales
- Рід Fragilaria
- Рід Asterionella
- Рід Diatoma
Порядок Cymbellales
- Рід Rhoicosphenia
- Рід Gomphonema
- Рід Cymbella
Порядок Achnanthales
- Рід Planothidium
- Рід Cocconeis
Порядок Naviculales
- Рід Navicula
- Рід Pinnularia
Порядок Bacillariales
- Рід Nitzschia
Порядок Surirellales
- Рід Surirella
- Рід Cymatopleura
Клас Pedinellophyceae
- Рід Pedinella
Див. також
Література
- Ботаніка. Водорості та гриби : навч. посіб. / І. Ю. Костіков [та ін.]; за ред. І. Ю. Костікова, В. В. Джаган ; Київський нац. ун-т імені Тараса Шевченка. - 2-ге вид., перероб. - К. : Арістей, 2007. - 476 с. : рис. - ISBN 966-8458-67-2
- Ботаніка з основами гідроботаніки (водні рослини України) / Б.Є. Якубенко, П.М. Царенко, І.М. Алейніков, С.І. Шабарова, С.П. Машковська, Л.М. Дядюша, А.П. Тертишний. – К.: Фітосоціоцентр, 2014 – 444 с.
- Вища водна рослинність / Д. В. Дубина ; відп. ред. Ю. Р. Шеляг-Сосонко. – К.: Фітосоціоцентр, 2006. – 412 с. - (Рослинність України)
- Водорості в системі органічного світу / Н. П. Масюк, І. Ю. Костіков ; НАН України, Ін-т ботаніки ім. М. Г. Холодного, Київ. нац. ун-т ім. Тараса Шевченка. - К. : Академперіодика, 2002. - 178 с. - Бібліогр.: с. 115-141. - ISBN 966-8002-38-5 (DjVu-файл)
- Водорості грунтів України : історія та методи дослідження, система, конспект флори / І. Ю. Костіков [та ін.] ; відп. ред. Н. П. Масюк, С. Я. Кондратюк ; Київ. нац. ун-т ім. Тараса Шевченка, Міжнар. Соломонів ун-т, Ін-т ботаніки ім. М. Г. Холодного НАН України, Мелітоп. пед. ун-т. - К. : Фітосоціоцентр, 2001. - 299 с. - Бібліогр.: с. 258-283. - ISBN 966-7938-40-9 (DjVu-файл на twirpx.com)
Джерела
- Дігтяр С.В. Розробка біотехнології переробки масових форм гідробіонтів : Рукопис // Одеська національна академія харчових технологій Міністерства освіти і науки України : Дисертація на здобуття наукового ступеня кандидата технічних наук за спеціальністю 03.00.20 – Біотехнологія. — Одеса, 2019.
- Ботаніка. Водорості та гриби: Навчальний посібник, 2-ге видання, переробл. – К.: Арістей, 2006. – 476 с.
- ВОДОРОСТІ // ЕСУ
- Белякова Г.А. и др. - Ботаника в 4 т., т.1, Водоросли и грибы.