Гриби
Гриби́ — гетерогенна група еукаріотичних безхлорофільних гетеротрофних організмів, які живляться переважно осмотрофно, і більшість з яких здатна розмножуватись за допомогою спор (хоча деякі втратили цю можливість і розмножуються вегетативно). Більшість з них протягом усього життя або на певних стадіях розвитку має міцеліальну будову, а деякі — дріжджі — одноклітинні. Наука про гриби — мікологія.
На початку XXI сторіччя було описано приблизно 70—100 тис. видів грибів[2][3], проте їх очікуване різноманіття, за оцінками різних авторів, становить від 300 тис. до 1,5 млн видів[3][4][5].
? ГрибиЧас існування: Раній Девон–наш час 410–0 Ma | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 За годинниковою стрілкою починаючи зверху зліва: мухомор (Amanita muscaria), саркосціфа (Sarcoscypha coccinea), різопус чорніючий (Rhisopus nigricans), хітридіомікотовий гриб (Chytridiomycota), пеніциліум (Penicillium) | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
Підцарства/Відділи/Підвідділи | ||||||||||||||||
| ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||
Хоча гриби переважно ростуть у ґрунтах, вони також поширені в більшості типів біотопів — морях, континентальних водоймах. Вони розвиваються на різноманітних природних субстратах рослинного та тваринного походження, на штучних матеріалах, створених людиною. Серед грибів відомі сапротрофи, симбіонти й паразити рослин та тварин, зокрема людини. Плодові тіла (спорокарпи) деяких грибів вживають в їжу: білий гриб(Boletus edulis Bull. ех Fr.), печериця(Agaricus), лисички(Cantharellus cibarius Fr.), сироїжка(Russula atrorubens), грифола(Grifola frondosa), трюфель(Tuber) тощо.
У побуті грибом часто називають спорокарп (плодове тіло) вищих грибів, що має ніжку і спороносну шапку з радіальними пластинами чи трубками. Також у побуті вживають назву грибок для позначення мікроскопічних пліснявих грибів. Обидві ці назви некоректні з наукового погляду й не відповідають науковому визначенню цієї групи біоти.
Відмінність грибів від рослин і тварин





Від рослин гриби відрізняються насамперед відсутністю фотосинтетичного апарата, наявністю хітинової клітинної стінки, утворенням сечовини в процесі метаболізму, тваринним шляхом синтезу амінокислот, запасанням глікогену тощо; від тварин — переважанням осмотрофного типу живлення над фаготрофним, розмноженням за допомогою спор та наявністю клітинних стінок. Проте ці відмінності грибів від тварин не є універсальними. Наприклад, до організмів, які традиційно (зокрема в Міжнародному кодексі ботанічної номенклатури) розглядають як гриби, належать міксомікотові слизовики. Вони позбавлені клітинних стінок і здатні до фаготрофного живлення (хоча розмножуються за допомогою спор).
Систематика
Мікологія традиційно була підрозділом ботаніки. Гриби, як окрему групу, виділили за їх морфологічною подібністю та особливостями живлення (насамперед за відсутністю фотосинтезу)[6]. Наприкінці 1960-х років Роберт Віттекер запропонував виділити гриби в окреме царство.
Однак у 1970—80-х роках у практику систематичних досліджень було впроваджено методи порівняльної цитології, а в 1990-х роках — методи молекулярної таксономії. Це призвело до революційної зміни уявлень про систематику еукаріот і, зокрема, про місце в цій системі грибів та грибоподібних організмів[3]. За образним висловлюванням, «стало зрозуміло, що слизовики мають такий же стосунок до грибів, як і кити — до риб»[7].
На початку XXI сторіччя грибів, яких історично поділяли на 10 відділів, об'єднали у три великі підгрупи, що належали до різних біологічних царств[3][8]:
- Справжні гриби, які розглядають як самостійне підцарство Fungi:
- Хітридіомікотові гриби [Chytridiomycota]- клас грибів відділу Хітридіомікотові гриби (Chytridiomycota).
- Зигомікотові гриби [Zygomycota]— тип грибів, що об'єднує 10 порядків, 27 родин, близько 170 родів і понад 1000 видів.
- Аскомікотові гриби [Ascomycota], сумчасті гриби – відділ справжніх грибів, у яких вегетативне тіло має вигляд розгалуженого септованого міцелію, рідше – плазмодія, дріжджового талому і псевдоміцелію.
- Базидієві гриби [Basidiomycota].
- Псевдогриби — споріднені з фотосинтезуючими організмами, однак вторинно втратили пластиди; підцарство страменопілів:
- Оомікотові [Oomycota], клас еукаріотичних осмотрофних організмів, у яких клітини вкриті целюлозно-глюкановою оболонкою, а вегетативне тіло представлено багатоядерним розгалуженим неклітинним міцелієм. Спочатку класифікувалися як гриби (звідки й походить «-міцети» чи «-мікоти» у назві), наприкінці 1990-х перекласифіковані до гетероконтів.
- Лабіринтуломікотові [Labyrinthulomycota]- відділ псевдогрибів. Осмотрофні страменопіли, у яких голі клітини вкриті мікроскопічними лусочками. Вегетативне тіло представлено поодинокими або з'єднаними у сітчасті псевдоплазмодії клітинами з ектоплазматичним ретикулюмом, який утворюють спеціальні органели — ботросоми. Монадні стадії дводжгутикові. Відділ включає близько 50-и видів псевдогрибів, поширених переважно у морських біотопах.-
- Гіфохітриомікотові [Hyphochytriomycota];— евкаріотичні осмотрофні страменопіли, у яких клітини у дорослому стані повністю або частково вкриті целюлозно-хітиновою оболонкою, а вегетативне тіло представлено поодинокими багатоядерними клітинами або ризоміцелієм.
- Слизовики:
- Акразіомікотові слизовики [Acrasiomycota] — підцарство дискокристат, відділ грибоподібних організмів, евкаріотичні первинно-гетеротрофні дискокристати, у яких клітини голі, а вегетативне тіло представлено амебоїдами (міксамебами). Відділ нараховує менше 20 видів мікроскопічних амебоїдних організмів.
- Міксомікотові слизовики (Myxomycota) — підцарство амебозоїв;група найпростіших клади Amoebozoa, що за зовнішнім виглядом, наявністю плодових тіл, та способом життя нагадують гриби, проте відрізняються клітинною організацією, особливостями розмноження та життєвого циклу і не споріднені із ними філогенетично. Первинно-гетеротрофні тубулокристати.
- Плазмодіофоромікотові слизовики (Plasmodiophoromycota) — підцарство ризаріїв;
Несистематизовану групу становили анаморфні (Deuteromycetes) і ліхенізовані гриби або лишайники (Lichenes)[джерело?]
Слизовиків та псевдогриби стали об'єднувати під загальною назвою «грибоподібні організми»[3], на відміну від «справжніх грибів».
Справжні гриби
До справжніх грибів традиційно включали чотири відділи: хітридіомікотові (Chytridiomycota), зигомікотові (Zygomycota), аскомікотові (Ascomycota) та базидієві гриби (Basidiomycota)[6]. Біохімічні та цитологічні дані, отримані у 1970—1980 роках (шляхи біосинтезу, структура клітинної стінки, особливості органел), свідчили на користь їх спорідненості. Молекулярно-генетичні дані 1990-х років підтвердили виокремлення такої групи, що отримала назву Fungi. Дані філогенетичного аналізу також засвідчили, що відділи міксомікотових, гіфохітриомікотових та оомікотових), яких раніше класифікували «грибами», до виділеної групи не належать[6].
Водночас філогенетичні дані показали близьку спорідненість із виділеною групою деяких одноклітинних організмів (зокрема, мікроспоридій, криптоміцетів та Pneumocystis), яких раніше класифікували до протистів[6]. Разом зі справжніми грибами (Fungi) їх об'єднали в кладу Holomycota [=Nucletmycea].
Філогенетичні дані наводять на думку, що справжні гриби (Fungi) набагато ближчі до тварин, аніж до рослин[6].
Кладу Holomycota разом із сестринською кладою Holozoa (до якої входять тварини), об'єднали до клади опістоконтів.
Найдавніші палеонтологічні знахідки слідів грибів виявлено у вулканічних породах віком 2,4 млрд років, щоправда, організми поселилися в них набагато пізніше. За будовою вони нагадують грибний міцелій ниткоподібної будови.[9][10]
Псевдогриби
До псевдогрибів («несправжніх грибів») об'єднали три відділи[11]:
- Оомікотові (Oomycota),клас еукаріотичних осмотрофних організмів, у яких клітини вкриті целюлозно-глюкановою оболонкою, а вегетативне тіло представлено багатоядерним розгалуженим неклітинним міцелієм. Спочатку класифікувалися як гриби (звідки й походить «-міцети» чи «-мікоти» у назві), наприкінці 1990-х перекласифіковані до гетероконтів.
- Лабіринтуломікотові (Labyrinthulomycota [=Labyrinthista]) відділ псевдогрибів. Осмотрофні страменопіли, у яких голі клітини вкриті мікроскопічними лусочками. Вегетативне тіло представлено поодинокими або з'єднаними у сітчасті псевдоплазмодії клітинами з ектоплазматичним ретикулюмом, який утворюють спеціальні органели — ботросоми. Монадні стадії дводжгутикові. Відділ включає близько 50-и видів псевдогрибів, поширених переважно у морських біотопах.
- Гіфохітриомікотові (Hyphochytriomycota); евкаріотичні осмотрофні страменопіли, у яких клітини у дорослому стані повністю або частково вкриті целюлозно-хітиновою оболонкою, а вегетативне тіло представлено поодинокими багатоядерними клітинами або ризоміцелієм.
Усіх їх класифікували до типу гетероконтів (Heterokonta [=Stramenopiles]).
Спочатку гетероконтів включили до царства хромістів (Chromista)[11], потім — до хромальвеолятів (Chromalveolata) і, врешті-решт (під назвою страменопіли) — до супергрупи SAR.
Слизовики
Систематика слизовиків виявилася найскладнішою. Група є вочевидь поліфілетичною і на початку 2000-х її класифікацію неодноразово переглядали. Станом на 2008 рік вважалося, що слизовики належать до п'яти окремих ліній[11]:
- Ramicristates [=Mycetozoa], до якої належали Dictyostelea [=Dictyosteliomycetes], Myxogastrea [=Myxomycetes], Protostelea [=Protosteliomycetes]
- три лінії (які раніше вважали однією) у складі акразіомікотовіх:
- Acrasiomycetes [=Heterolobosea]
- Copromyxida [=Copromyxa]
- Fonticulida [=Fonticula]
- Плазмодіофоральні (Plasmodiophorida [=Plasmodiophorea] [=Plasmodiophoromycetes]) — включені до Cercozoa.
Біохімічні ознаки
Біосинтез лізину
Амінокислота лізин належить до незамінних, багатоклітинні тварини її не синтезують. В еукаріотичному світі відомо два основних шляхи біосинтезу лізину: через α-аміноадипінову кислоту (т. зв. ААА-шлях) та через диамінопімелінову кислоту (ДАП-шлях).
Лізин за ААА-шляхом синтезується з ацетилкофермента А та α-кетоглутарової кислоти. Вони утворюють α-аміноадипінову кислоту, що надалі перетворюється на лізин. Цей шлях також називають грибним шляхом біосинтезу, оскільки за ним лізин синтезують справжні гриби (відділи Chytridiomycota, Zygomycota, Ascomycota та Basidiomycota), хоча цей шлях біосинтезу притаманний також евгленофітовим водоростям (вони ж Евгленові).
Синтез за ДАП-шляхом відбувається з аспартату та пірувату через диамінопімелінову кислоту. Цей шлях біосинтезу називають також рослинним, оскільки саме за ним відбувається біосинтез лізину у всіх еукаріотичних водоростей (крім евгленофітових) та вищих рослин, а також у псевдогрибів з відділів Oomycota та Hyphochytriomycota.
Для частини грибів лізин є незамінною амінокислотою, яку організм сам не здатний синтезувати. Зокрема, це спостерігають у псевдогрибів з Labyrintulomycota.
Біосинтез триптофану
Амінокислоту триптофан гриби синтезують в результаті роботи специфічних груп ферментів. Загалом у грибів відомо п'ять таких груп ферментів. Перші чотири групи притаманні грибам — платикристатам, п'ята група — оомікотовим грибам. Ферментні комплекси першої групи виявлено у плазмодіофоромікотових, хітридіомікотових та аскомікотових грибів. Друга група — у шапинкових базидієвих грибів. Третя група характерна для дріжджоподібних аскомікотових, а четверта — для зигомікотових та базидієвих грибів (крім шапинкових).
Утворення сидерамінів
Сидерамінами називають складні органічні сполуки, які транспортують у клітину залізо. Сидераміни синтезують всі справжні гриби, крім зигомікотових. Далі молекули цих речовин виділяються у зовнішнє середовище, зв'язуються з іонами заліза, і знов поглинаються клітиною гриба. Псевдогриби (зокрема, оомікотові) сидераміни не утворюють, і поглинання іонів заліза в них здійснюється іншими способами.
Продукти асиміляції
Основними продуктами асиміляції майже в усіх грибів є β-1,4-глюкани, які представлені тваринним глікогеном. У акразіомікотових слизовиків основним продуктом асиміляції є близький до парамілону β-1,3-глюкан або β-1,6-глюкан. У оомікотових грибів основний продукт асиміляції — це також β-1,3-глюкан, який, проте, близький до ламінарину та хризоламінарину, і має назву міколамінарин. Найпоширенішим додатковим асимілятом виступає олія, яку запасають у великих кількостях переважно спори та старіючі клітини.
Способи живлення грибів
Всі гриби є облігатно гетеротрофними організмами. Проте поглинання органічних речовин вони можуть здійснювати двома шляхами — осмотрофно (піноцитозом) та фаготрофно (фагоцитозом).
При осмотрофному живленні організм або його окремі клітини (наприклад, клітини трофічних гіф — гаусторій) поглинають розчинену органічну речовину шляхом абсорбції, без утворення травних вакуолей. Зазвичай абсорбуються низькомолекулярні органічні речовини, які утворюються при розщепленні високомолекулярних сполук різноманітними гідролітичними екзоферментами. Грибні екзоферменти здатні розкладати до мономерів такі біополімери як целюлозу, лігнін, хітин, білки, нуклеїнові кислоти тощо.
При фаготрофному живленні органічну речовину організм поглинає у вигляді твердих часток за допомогою псевдоподій, і надалі перетравлює або у травних вакуолях, або в лізосомах. Осмотрофний тип живлення притаманний всім грибам, тоді як фаготрофний — лише міксомікотовим слизовикам.
Відомі також хижі гриби, що полюють на нематод або паразитують на них, наприклад Arthrobotrys oligospora.[12]
Будова клітини
Клітинні покриви



У справжніх грибів та грибоподібних організмів клітини можуть бути:
- голими;
- вкритими ектоплазматичним ретикулюмом (саме зовнішній, а не внутрішній — ред.);
- мати клітинну оболонку.
Голі клітини, які вкриті лише плазмалемою, зазвичай здатні до амебоїдного руху та фаготрофного живлення. Відсутність клітинних покривів характерна для вегетативних тіл акразіомікотових, міксомікотових та плазмодіофоромікотових слизовиків та для частини справжніх грибів з відділу хітридіомікотових. Голі клітини мають також зооспори псевдогрибів та справжніх грибів.
У плазмодіофоромікотових слизовиків вегетативне тіло, представлене багатоядерним плазмодієм, перед початком утворення спор вкривається тонкою оболонкою нез'ясованої хімічної природи, розташованою назовні від плазмалеми. Цей покрив помітний лише при електронній мікроскопії. Плазмодії плазмодіофоромікотових, хоча і є голими, не здатні до фаготрофного живлення і не утворюють псевдоподій.
У лабіринтуломікотових псевдогрибів над плазмалемою розташовується ектоплазматичний ретикулюм. Він являє собою додатковий зовнішній одномембранний покрив, що утворює численні довгі тонкі випини, які анастомозують між собою та з ектоплазматичним ретикулюмом інших клітин, утворюючи складну сітчасту структуру. Простір між мембраною ектоплазматичного ретикулюму та плазмалемою виповнений однорідним аморфним матриксом. Мембрани ектоплазматичного ретикулюму продукують особливі одномембранні органели — ботросоми (сагеногеносоми). Крім того, на поверхні клітин лабіринтуломікотових виявлено субмікроскопічні лусочки, які утворені у везикулах, похідних від комплексу Гольджі.
Найпоширенішим типом клітинних покривів, який притаманний більшості псевдогрибів та справжніх грибів, є клітинна оболонка. У грибів вона здійснює не лише захисну, але й ряд інших важливих функцій, зокрема, є місцем локалізації гідролітичних екзоферментів, бере участь в абсорбції поживних речовин із субстрату, в морфогенетичних та ростових процесах, надає форму клітинам грибних гіф та органам розмноження. Зовні клітинна оболонка може бути вкрита шаром слизу, який утворює капсулу.
Основу клітинної оболонки складають мікрофібрилярні скелетні компоненти — хітин або целюлоза. Над ними розташовується зовнішній аморфний матрикс, утворений глюканами, хітозаном або маннаном. З мікрофібрилярними та аморфними компонентами пов'язані різні білки та ліпіди, а також пігменти (меланіни, хінони), розчинні цукри, амінокислоти, різноманітні іони та солі.
В межах конкретних відділів грибів хімічний склад оболонки вважають консервативною ознакою високої таксономічної ваги. Зокрема, скелетним компонентом оболонок псевдогрибів є целюлоза, а справжніх грибів — хітин. Структурні компоненти аморфного матриксу у різних відділах кожної з цих груп різні.
Загалом за хімічним складом мікрофібрилярного та аморфного компонентів оболонки грибів поділяють на целюлозно-глюканові (Oomycota), целюлозно-хітинові (Hyphochytriomycota), хітин-глюканові (Chytridiomycota, Basidiomycota), хітин-глюканові, інколи — з мананами (Ascomycota) та хітин-хітозанові (Zygomycota). Проте відомо також ряд винятків з правила сталості типу клітинної оболонки в межах відділу. Зокрема, у дріжджів, які належать до аскомікотових грибів, в оболонках може бути відсутній хітин, а у представників порядку моноблефаридальних з відділу хітридіомікотових виявлено целюлозу. У слизовиків, вегетативні стадії яких позбавлені клітинних покривів, оболонками вкриті спори. Зокрема, у спор слизовиків з Myxomycota та представників порядку Acrasiales з Acrasiomycota основу оболонки складає целюлоза, а у слизовиків з Plasmodiophoromycota — хітин.
Ядерний апарат
Ядро клітини грибів має типову для всіх еукаріотів будову — воно оточене двомембранною ядерною оболонкою, містить одне або кілька ядерець; ядерна ДНК пов'язана з гістонами і має хромосомну організацію. Різноманітність ядерного апарату у грибів різних відділів проявляється, в першу чергу, у кількості ядер в клітині та в особливостях мітозу.
Ядерний геном. У грибів ядерний геном має проміжний розмір між геномом бактерій й вищих еукаріот. Наприклад, пекарські дріжджі мають 15 хромосом, але кожна хромосома у середньому у 5 разів менша хромосоми кишкової палички й у 4 рази більша ДНК Т-фагу. У середньому розмір геному у грибів на два порядки менший, ніж у вищих рослин. Кількість хромосом різних грибів становить 2-28, але більшість видів має 10-12 хромосом. Більші коливання відзначені у розмірах ДНК на гаплоїдний геном: від 0,015 нг у дріжджів Saccharomyces до 8,2 нг у зігомецету Entomophaga, тобто, вміст ДНК відрізняється більш як у 500 разів (у вищих рослин менш ніж у 100 разів). Найменші розміри геному має аскоміцет Aeshbya gossypii — 9,7 млн пар нуклеотидів (у цукроміцетних дріжджів — 13,5 млн, у більшості грибів — 25—40 млн). Це найменший геном серед усіх досліджених еукаріот (менші розміри ДНК мають лише нуклеоморфи деяких водоростей, але вони перебувають в ендосимбіотичних структурах, а не у вільноживучих організмах).
Структура ядерного геному грибів також проміжна між бактеріальним й геномом вищих еукаріот. У істинних грибів процент повторюваних послідовностей низький (10-15 %); у бактерій повтори майже відсутні, а у вищих еукаріот складають значну частину геному. Повтори у геномі грибів представлені майже винятково рибосомальними генами. Виняток складають ооміцети (псевдогриби), у геномі яких повтори складають 18-65 %. Грибні інтрони (некодовані послідовності ДНК, які вирізаються перед трансляцією) також дрібніші, ніж у вищих еукаріот. Таким чином, частка ДНК, яка бере участь у синтезі білків, у грибів більша, ніж у вищих еукаріот.
Також у багатьох грибів виявлено B-хромосоми. Інформація, яка закодована у них, не є обов'язковою для життєзабезпечення клітин, але іноді вона важлива для адаптації до умов середовища[13].
Кількість ядер. Вегетативні тіла грибів можуть бути одно- та багатоклітинними, або мати неклітинну будову. Клітини одно- та багатоклітинних грибів зазвичай або одноядерні або містять два генетично неоднакових ядра, які називають дикаріоном. Дикаріони є асоціацією двох ядер, що утворилися після злиття цитоплазм двох клітин без подальшої каріогамії. Одноядерні та дикаріонтичні клітини характерні для вегетативних стадій лабіринтуломікотових, аскомікотових та базидіомікотових грибів. Крім того, одноядерними є вегетативні амебоїдні клітини акразіомікотових слизовиків, а також спори більшості грибів. Неклітинний план будови тіла, при якому одна клітина містить багато ядер, відомий у кількох відділах слизовиків, псевдогрибів та справжніх грибів. Багатоядерні клітини за ступенем генетичної неоднорідності ядер поділяють на три групи:
- ті, що містять лише генетично однакові ядра;
- ті, що містять багато пар дикаріонів, тобто є полідикаріонтичними;
- ті, що містять багато генетично відмінних ядер, тобто гетерокаріонтичні.
Багатоядерні клітини з генетично однаковими ядрами утворюються тоді, коли багатоядерність виникає внаслідок серії мітозів лише одного вихідного ядра (наприклад, ядра спори). Такі вегетативні клітини мають оомікотові, гіфохітриомікотові, хітридіомікотові, зигомікотові та деякі аскомікотові гриби.
Клітини з багатьма дикаріонами утворюються тоді, коли при статевому процесі після плазмагамії каріогамія одразу не відбувається, а ядра щойно утвореного дикаріону починають синхронно мітотично ділитися. Саме такий тип клітин мають вторинні плазмодії плазмодіофоромікотових слизовиків. Первинні плазмодії представників цього відділу утворюються внаслідок серії послідовних мітозів ядра однієї зооспори, і, таким чином, всі ядра в первинному плазмодії, на відміну від вторинного, є генетично однаковими.
Гетерокаріонтичність характерна для плазмодіїв більшості міксомікотових слизовиків, оскільки багатоядерність у цих організмів розвивається не лише внаслідок багатьох мітозів, але й злиття з цитоплазмами інших (часто — багатьох) одно- та багатоядерних особин.
Різноманітність мітозу у грибів проявляється в першу чергу в наявності центріолей, поведінці ядерної оболонки та ядерця. У акразіомікотових та плазмодіофоромікотових слизовиків, всіх псевдогрибів, а також хітридіомікотових грибів, мітоз відбувається за участю центріолей. Цікаво, що представники цих відділів в життєвому циклі мають репродуктивні джгутикові стадії. У тих відділах справжніх грибів, де монадні стадії відсутні (у Zygomycota, Ascomycota та Basidiomycota), відсутні також і центріолі. У слизовиків з Myxomycota центріолі здебільшого є, але вони стають центрами організації веретена поділу і беруть участь у мітозі лише під час поділу одноядерних клітин — зооспор чи міксамеб; у багатоядерних плазмодіях мітоз відбувається без участі центріолей, навіть коли вони є. Більшість грибів під час мітозу зберігають ядерну оболонку. У акразіомікотових, міксомікотових, оомікотових, зигомікотових та аскомікотових грибів вона залишається інтактною, і мітоз, таким чином, є закритим. Крім того, на зовнішній ядерній мембрані у зигомікотових грибів у метафазі з'являється специфічна дископодібна структура. Плазмодіофоромікотові, лабіринтуломікотові, гіфохітриоміктові та хітридіомікотові біля кожного полюсу веретена в ядерній оболонці утворюють по одній великій перфорації, тобто, мітоз є напівзакритим. Відкритий мітоз виявлено лише у міксомікотових слизовиків (на стадії одноядерних міксамеб) та у деяких шапинкових базидієвих грибів. Ядерце у грибів при мітозі має три варіанти поведінки. У першому варіанті воно зникає на початку мітозу і відновлюється після його закінчення (Myxomycota, Labyrintulomycota, Hyphochytriomycota). У другому варіанті ядерце не зникає, і на певній стадії ділиться перешнуровкою: у профазі — у акразіомікотових, у метафазі — в плазмодіофоромікотових, на початку телофази — у оомікотових та більшості видів зигомікотових грибів. У частини видів з Chytridiomycota, Ascomycota та Basidiomycota спостерігається третій варіант поведінки ядерця: на початку мітозу воно виштовхується з ядра в цитоплазму, де й зберігається до закінчення поділу.
Мітохондріальний апарат
Всі гриби мають мітохондрії типової для решти еукаріотів будови — вони оточені двомембранною оболонкою, внутрішня мембрана утворює інвагінації — кристи. Матрикс мітохондрій містить мітохондріальну ДНК, прокаріотичні рибосоми з коефіцієнтом седиментації 70S, комплекс різноманітних ферментів, більшість з яких забезпечує процес дихання.
Різноманітність організацій мітохондріального апарату, як і у водоростей, простежують, в першу чергу, за морфологією крист. Зокрема, акразіомікотові слизовики, подібно до евгленофітових водоростей, мають кристи трьох типів — дископодібні, трубчасті та пластинчасті. Міксомікотові слизовики та псевдогриби містять мітохондрії з трубчастими та пластинчастими кристами або виключно з трубчастими. У плазмодіофоромікотових слизовиків та всіх справжніх грибів (Chytridiomycota, Zygomycota, Ascomycota, Basidiomycota) кристи лише пластинчасті.
Джгутикові стадії
Представники всіх відділів слизовиків, псевдогрибів та одного з відділів справжніх грибів (Chytridiomycota) в життєвому циклі здатні утворювати джгутикові стадії, представлені зооспорами або гаметами. У справжніх грибів з відділів Zygomycota, Ascomycota та Basidiomycota відсутні не лише джгутикові стадії, але й центріолі — органели, похідні від базальних тіл джгутиків.
У найзагальнішому вигляді джгутикові стадії грибів поділяють на три основні типи:
- двома ізо- або гетероконтними передніми гладенькими джгутиками (інколи один із джгутиків може бути редукованим, але зберігається його базальне тіло);
- з двома гетероконтними передніми або бічними джгутиками, з яких довший вкритий тричленними волосками — ретронемами (другий джгутик може бути редукованим до базального тіла);
- з одним заднім бичоподібним гладеньким джгутиком.
Джгутики першого типу характерні для всіх слизовиків. При цьому акразіомікотові мають монадні клітини з двома ізоконтними джгутиками, міксомікотові — з двома гетероконтними, плазмодіофоромікотові — з двома гетероконтними або одним джгутиком.
Монадні клітини другого, страменопільного плану будови, характерні для псевдогрибів. Утворення ретронем, які вкривають один із джгутиків, починається в міжмембранному просторі ядерної оболонки. Далі від зовнішньої мембрани ядра у цитоплазму відшнуровуються пухирці з зачатковими ретронемами. Після дозрівання везикули рухаються до поверхні клітини, зливаються з плазмалемою біля базальних тіл джгутиків і виводять, таким чином, ретронеми на плазмалему при основі аксонеми джгутика. Монадні клітини у різних відділах псевдогрибів відрізняють, в першу чергу, за кількістю джгутиків: у оомікотових та лабіринтуломікотових монадні стадії дводжгутикові, довший джгутик, вкритий ретронемами і направлений вперед; коротший гладенький, і орієнтований назад. Монадні клітини гіфохітриомікотових мають лише один передній пірчастий джгутик; від другого джгутика залишається тільки базальне тіло, видозмінене в поодиноку центріолю.
Монадні стадії третього типу — з гладким заднім бичоподібним джгутиком — називають інколи опістоконтними. Ультратонка будова таких клітин дуже схожа з будовою сперматозоїдів багатоклітинних тварин. Вважають, що опістоконтні клітини виникли від дводжгутикових внаслідок редукції одного з джгутиків. Це підтверджується наявністю у клітині поодинокої центріолі, розташованої поблизу базального тіла джгутика, яка, ймовірно, є видозміненим базальним тілом другого джгутика. Опістоконтні джгутикові стадії у грибів представлені лише у відділі Chytridiomycota, і розглядаються як один із доказів походження справжніх грибів та багатоклітинних тварин від спільного пращура.
Комплекс Гольджі
Комплекс Гольджі розвинений не у всіх грибів. Зокрема, його немає у акразіомікотових слизовиків та справжніх грибів (хітридіомікотових, зигомікотових, аскоміктових та базидіомікотових). У аскомікотових та базидіомікотових в клітинах замість комплексу Гольджі інколи спостерігають цистерни ендоплазматичної сітки, що утворюють невеликі стопки. Проте такі комплекси цистерн обов'язково асоціюються принаймні з одним з каналів ендоплазматичної сітки. Добре розвинені типові комплекси Гольджі наявні у плазмодіофоромікотових слизовиків та псевдогрибів (оомікотових, гіфохітриомікотових та лабіринтуломікотових). Зазвичай у цих грибів комплекс Гольджі притиснутий до поверхні ядерної мембрани.
Морфологія вегетативного тіла
Амебоїдна будова

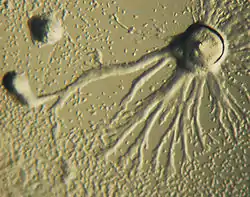
Амебоїні таломи позбавлені твердих клітинних покривів, здатні до метаболічних змін форми, можуть утворювати справжні або несправжні псевдоподії. Амебоїдні таломи характерні для всіх слизовиків, лабіринтуломікотових та гіфохітриомікотових та багатьох хітридіомікотових грибів. Залежно від кількості ядер амебоїдні вегетативні тіла поділяють на міксамеби, міксофлагеляти, плазмодії та псевдоплазмодії.
- Міксамеби завжди мікроскопічні, одноядерні або зрідка дикаріонтичні, позбавлені джгутиків, здатні до активного руху за допомогою псевдоподій. За відносними розмірами та кількістю псевдоподії міксамеб поділяють на видовжені вузькі та зазвичай численні філоподії та короткі широкі поодинокі лобоподії. Філоподії здатні захоплювати тверді частки їжі, тобто крім локомоторної виконують ще й трофічну функцію. Міксамеби з філоподіями притаманні міксомікотовим слизовакам. Міксамеби з поодинокою лобоподією, яка здійснює лише локомоторну функцію, характерні для акразіомікотових слизовиків. Лобоподія у міксамеб акразіомікотових розташовується на передньому кінці клітини. Задня частина міксамеби утворює лопать, в якій розташовуються видільні вакуолі, і називається уроїдною зоною.
- Міксофлагеляти також мікроскопічні, утворюють філоподії, але крім того мають два гетероконтних джгутики, і, таким чином, являють особливий варіант монадних клітин. Залежно від наявності крапельно-рідкої води та концентрації іонів кальцію міксофлагеляти здатні перетворюватись на міксамеби, і навпаки. Серед грибів вегетативні клітини, представлені міксофлагелятами, зустрічаються лише у відділі Myxomycota.
- Псевдоплазмодії являють собою агрегати міксамеб, які зберігають свою індивідуальність. Утворюються псевдоплазмодії внаслідок об'єднання у колонії поодиноких міксамеб, при цьому цитоплазми останніх між собою не зливаються. Псевдоплазмодії відомі у акразіомікотових слизовиків, лабіринтуломікотових псевдогрибів та міксомікотових слизовиків з класу диктіостеліоміцетів. Псевдоплазмодії кожного з цих таксонів відрізняють за способом агрегації клітинних індивідів, їх здатністю до руху взагалі та скоординованого руху зокрема.
- Плазмодії мають неклітинну будову: вони схожі на міксамеби, але містять багато ядер — від чотирьох до десятків та навіть сотень тисяч. Розміри плазмодіїв коливаються в широких межах — від мікроскопічних (наприклад, у плазмодіофоромікотових слизовиків) до макроскопічних (у багатьох міксомікотових). Плазмодії можуть бути нерухомі або здатними рухатись за допомогою псевдоподій. Відомо два способи утворення плазмодіїв: по-перше, з одноядерних міксамеб або репродуктивних клітин (зокрема, зооспор) внаслідок серії послідовних мітотичних поділів ядра; по-друге, внаслідок злиття між собою кількох (інколи — багатьох) амебоїдних одно- або багатоядерних індивідів. Плазмодії можуть також утворюватись внаслідок комбінації обох цих шляхів.
Зі способом утворення плазмодію пов'язаний ступінь генетичної однорідності його ядер, згідно з чим плазмодії поділяють на генетично однорідні, полідикаріонтичні та гетеродикаріонтичні. За розмірами, формою та наявністю в плазмодіях закономірного руху цитоплазми (т. зв. човникових рухів), плазмодії поділяють на протоплазмодії, афаноплазмодії та фанероплазмодії.
- Протоплазмодії мікроскопічні, зазвичай без певної форми або неправильно сітчасті, без човникових рухів цитоплазми. Такий тип плазмодіїв притаманний деяким міксомікотовим, плазмодіофоромікотовим слизовикам та частині хітридіомікотових грибів.
- Афаноплазмодії макроскопічні, неправильно-віялоподібної форми, з цитоплазмою, що здійснює човникові рухи. Афаноплазмодії зовні нагадують губчасту протоплазматичну масу. Характерні для частини міксомікотових слизовиків.
- Фанероплазмодії також макроскопічні. На відміну від афаноплазмодіїв вони мають досить щільну консистенцію і майже правильну віялоподібну форму. У фанероплазмодіях є система розгалужених жилок, по яких цитоплазма здійснює човникові рухи. Зовні фанероплазмодії вкриті шаром ущільненого слизу. Як і афаноплазмодій, цей тип вегетативного тіла зустрічається лише у представників Myxomycota.
Особливим варіантом вегетативного тіла, який часто розглядають, як перехідний між амебоїдним та міцеліальним планами будови, є ризоміцелій. Ризоміцелій складається з багатоядерної вкритої клітинною оболонкою центральної частини, від якої відходять голі тонкі розгалужені ризоподії, що позбавлені ядер. Такий тип вегетативного тіла відомий у представників відділів Hyphochytriomycota та Chytridiomycota.
Міцеліальна будова


Вегетативне тіло переважної більшості грибів являє собою систему вкритих клітинними оболонками ниток, яка називається міцелій. Окрема нитка є елементарною одиницею міцелію і називається гіфою. Зрідка міцелій складає лише одна гіфа, тобто є нерозгалуженим (у деяких хітридіомікотових грибів). Проте у більшості випадків міцелій галузиться, і його складає велика кількість гіф.
Гіфи мають вигляд циліндричних трубок, діаметр яких коливається в межах 2-150 мкм, проте найчастіше становить 5-10 мкм. Гіфа здатна до необмеженого росту завдовжки, причому цей ріст завжди апікальний, і обумовлений, в першу чергу, роботою літичних пухирців та хітосом, або їх аналогів. За будовою гіфи поділяють на два типи — несептовані та септовані.
Несептовані гіфи багатоядерні та позбавлені поперечних перегородок, тобто мають неклітинну будову. Міцелій, який утворений несептованими гіфами, називають неклітинним міцелієм. Такий тип міцелію притаманний майже всім оомікотовим грибам, частині хітридіомікотових та більшості зигомікотових грибів.
Міцелій, який складається з гіф, що рівномірно поділені на клітини поперечними перегородками — септами, називають клітинним, або септованим міцелієм. Клітини гіф септованого міцелію можуть містити або лише одне ядро, або один дикаріон, або кілька ядер. Септи, які розмежовують сусідні вегетативні клітини гіфи, майже завжди мають поровий апарат, завдяки якому міцелій являє собою фізіологічно цілісний багатоклітинний організм. Структура порового апарату септ у грибів різних таксономічних груп різна. За будовою пор септи поділяють на:
- мікропорові,
- прості,
- доліпорові.
Мікропорові септи перфоровані багатьма дрібними порами. Септи такого типу зустрічаються зрідка, і відомі лише у деяких представників хітридіомікотових, зигомікотових та аскомікотових грибів.
Прості септи мають лише одну досить велику центральну пору. Септа у напрямку до пори потоншується. Пора в простій септі може бути відкрита (у сажкових грибів з Basidiomycota) або прикрита спеціальними структурами — дрібними вакуолями (наприклад, у іржастих базидієвих грибів) або тільцем Вороніна (у багатьох аскомікотових).
Доліпорові септи також мають лише одну велику центральну пору, проте, на відміну від простих септ, поперечна перегородка навколо пори потовщена. Пора в доліпорових септах може бути відкритою, закритою пробкою з аморфної електронно-щільної речовини або прикрита пористим мембранним ковпачком — парентосомою. Доліпорові септи з парентосомами притаманні більшості базидієвих грибів з макроскопічними плодовими тілами. Доліпорові септи без парентосом поширені переважно серед зигомікотових грибів із септованим міцелієм.
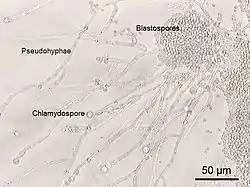
У деяких грибів вегетативне тіло має вигляд поодиноких клітин, що брунькуються. Якщо дочірні клітини після утворення септи не відокремлюються від материнської, то утворюється ланцюжок фізіологічно не пов'язаних між собою клітин, який називають псевдоміцелієм. Такий тип вегетативного тіла властивий деяким аскоміцетам, зокрема він характерний для дріжджів.
Розмноження
Вегетативне розмноження
Відбувається внаслідок поділу одноклітинних індивідів (міксамеб і міксофлагелят), фрагментацією плазмодіїв та міцелію. До способів вегетативного розмноження відносять також розмноження шляхом утворення артроспор та хламідоспор. Артроспори виникають внаслідок фрагментації гіфи на окремі короткі клітини. Хламідоспори утворюються подібно до артроспор, але, на відміну від останніх, мають темнозабарвлені, переважно потовщені клітинні оболонки, і є аналогом акінет у водоростей. Брунькування, яке супроводжується відокремленням дочірніх клітин, також вважають одним із способів вегетативного розмноження.
Нестатеве розмноження
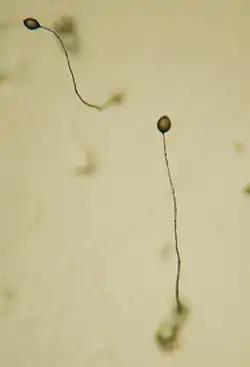
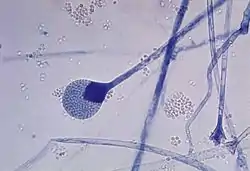
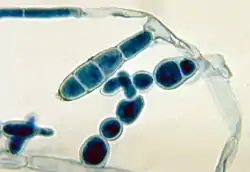
Здійснюють спеціалізовані клітини — спорами, які можуть утворюватись ендогенно у спорангіях та екзогенно, відокремлюючись від спеціалізованих гіф — конідієносців.
До спеціалізованих клітин нестатевого розмноження, що мають ендогенне походження, відносять спори міксомікотових слизовиків (за винятком диктіостеліоміцетових), міксамеби, зооспори, а також спорангіоспори зигомікотових грибів.
- Спори міксомікотових нерухомі, вкриті целюлозною оболонкою, і утворюються у спорангіях або всередині плодових тіл. У еугамних слизовиків утворенню спор передує редукційний поділ ядер; такі спори мають гаплоїдний набір хромосом. Залежно від наявності у середовищі крапельно-рідкої води спора проростає поодинокою міксамебою, міксофлагелятою або зооспорою.
- Міксамеби. Розмноження за допомогою міксамеб притаманне акразіомікотовим та міксомікотовим слизовикам. Міксамеби утворюються при проростанні ендогенних (у міксомікотових) або екзогенних (у акразіомікотових) спор.
- Зооспори відомі в усіх відділах слизовиків, псевдогрибів та у справжніх грибів з відділу хітридіомікотових. Зооспори грибів завжди позбавлені клітинної оболонки. Ультратонка будова зооспор є однією з найвагоміших таксономічних ознак, яку використовують при поділі грибів на відділи. В різних відділах грибів зооспори утворюють спорангії різних типів. Зокрема, у слизовиків зооспори розвиваються із ендогенних (міксомікотові) або екзогенних (акразіомікотові та плазмодіофоромікотові) нерухомих спор, причому з однієї спори виходить лише одна зооспора (фактично нерухома спора слизовиків являє собою спорангій з однією зооспорою або міксамебою). Зооспори лабіринтуломікотових утворюють соруси цист. Такі цисти виникають всередині однієї клітини після серії мітозів, які завершуються редукційним поділом. Одна циста дає початок одній зооспорі з гаплоїдним набором хромосом. У гіфохітриомікотових, оомікотових та хітридіомікотових грибів один зооспорангій утворює багато зооспор. Гриби, у яких на зооспорангій перетворюється все вегетативне тіло, називають холокарпічними. Представників, у яких спорангієм стає лише частина вегетативного тіла, належать до еукарпічних форм. Зокрема, всі гіфохітриоміцети є організмами холокарпічними. У хітридіомікотових відомі як холо-, так і еукарпічні представники.
- Спорангіоспори. Спорангії оомікотових здатні розвиватись або безпосередньо з верхівкової частини вегетативної гіфи, яка відокремлюється від решти міцелію септою (переважно у тих видів, що мешкають у водному середовищі), або відшнуровуватись від спеціалізованої гіфи — спорангіофора (у більшості наземних представників). Спорангій, який розвивається на спорангіофорі, є багатоядерним, і, залежно від умов зволоження, проростає або зооспорами, або амебоїдним протопластом, або неклітинною гіфою. У водних оомікотових зооспори здатні змінювати свою морфологію. Це явище отримало назву дипланетизму: зооспора, яка виходить зі спорангію, має апікальні джгутики; після деякого періоду активного руху вона інцистується; при проростанні зооспора виходить з оболонки цисти, але тепер її джгутики розташовуються латерально. Спорангіоспори також утворюються ендогенно, але, на відміну від зооспор, міксамеб та спор слизовиків, позбавлені джгутиків, вкриті міцною оболонкою, і проростають у гіфи міцелію. Спорангіоспори формують спорангії в своєму просторі. Ці спорангії розташовані на верхівках спеціалізованих гіф — спорангієносців, які піднімають органи нестатевого спороношення над субстратом.
- Спори екзогенного походження. До спеціалізованих клітин нестатевого розмноження, що мають екзогенне походження, відносять спори плазмодіофоромікотових та акразіомікотових слизовиків та конідії. Спори плазмодіофоромікотових та акразіомікотових слизовиків розвиваються внаслідок трансформації та фрагментації частин плазмодіїв та псевдоплазмодіїв без утворення спорангіїв. Подібно до спор міксомікотових, вони проростають поодинокими міксамебами або зооспорами.
- Конідії. Розмноження конідіями є основним і найпоширенішим способом нестатевої репродукції переважної більшості справжніх грибів. Конідії — це нерухомі, вкриті оболонкою клітини, що утворюються екзогенно, відокремлюючись від спеціалізованих гіф міцелію — конідієносців. На відміну від артроспор та хламідоспор, які проходять стадію диференціації лише після відокремлення від вегетативної гіфи, диференціація конідій розпочинається ще на міцелії: верхівкова частина конідієносця перетворюється на зачаток конідії, часто збільшується у розмірах, інколи синтезує додаткові шари оболонки, і далі відокремлюється від гіфи. Конідії, які спочатку відокремлюються від конідієносця септою, а далі дозрівають на ньому, називають алевроконідіями. Перед відокремленням від материнської гіфи пори, які з'єднували зачаткову алевроконідію з протопластом конідієносця, закривають пробки. Конідії, які спочатку проходять стадію диференціації, і лише потім відокремлюються септою від конідієносця, називають бластоконідіями. Конідієносці мають різноманітну форму — від поодиноких нерозгалужених до складно багаторазово розгалужених. Конідії на конідієносцях можуть бути поодинокими або утворювати довгі ланцюжки. По відношенню один до одного конідієносці можуть розташовуватись різними способами. За характером розташування конідієносців виділяють кілька типів конідіальних спороношень: поодинокі конідієносці, коремії, спородохії, ложа та пікніди. Поодинокі конідієносці щільних груп не утворюють. Якщо конідієносці розвиваються щільною групою, склеюючись боками за допомогою слизу і утворюючи колонку з головкою конідій на верхівці, то таке спороношення називають коремієм. Суцільні шари конідієносців, які розвиваються у вигляді подушечок на опуклій системі щільно переплетених гіф, утворюють спородохії. Ложа нагадують спородохії, але тут конідієносці розвиваються не на опуклому, а на плоскому переплетенні гіф. Якщо переплетення гіф, на якому розвиваються конідієносці, глибоко увігнуте і нагадує горщик, занурений у субстрат або у структуру з видозмінених вегетативних гіф (т. зв. строму), то таке спороношення називають пікнідою. Типи конідіальних спороношень є важливою ознакою в таксономії справжніх грибів на рівнях, починаючи з класів і закінчуючи видами. Проте особливого значення морфологія конідіальних спороношень набуває у таксономії т. зв. мітоспорових грибів — тих аскомікотових та базидієвих, у яких статеві спороношення не відомі, а місце у системі поки що є нез'ясованим. У справжніх грибів вегетативну стадію, яка утворює лише нестатеве (переважно — конідіальне) спороношення, називають анаморфною, на відміну від вегетативної стадії, яка бере участь у статевому процесі і утворюють статеві спороношення — т. зв. телеоморфної. Разом анаморфна та телеоморфна стадії складають повний цикл розвитку гриба — голоморфу.
Статеве розмноження
Відбувається внаслідок статевого процесу, який, як і у інших еукаріотів, обумовлює підтримання певного рівня рекомбінантної мінливості популяцій. Статевий процес серед грибів не виявлено лише у акразіомікотових слизовиків, і достовірно не описано у лабіринтуломікотових (хоча ультраструктурні дані свідчать про наявність в останньому відділі мейозу, який відбувається при утворенні цист). Як і у випадку нестатевого розмноження, статеві процеси у грибів різноманітніші, ніж у водоростей, і представлені гологамією, різними варіантами гаметогамії, гаметангіогамією та соматогамією.
Гологамія характерна для слизовиків з відділу Myxomycota, де відбувається внаслідок злиття вегетативних амебоїдів (переважно міксамеб).
Гаметогамія представлена ізо-, гетеро- та оогамією. Ізогамію виявлено у міксомікотових та плазмодіофоромікотових слизовиків, а також у гіфохитріомікотових псевдогрибів та більшості еугамних хітридіомікотових. Гетерогамію в цілому у грибів зустрічають зрідка, і відома вона лише у деяких хітридіомікотових.
- Оогамний статевий процес зустрічають у частини хітридіомікотових з порядку моноблефаридальних та у абсолютної більшості оомікотових (останній відділ отримав назву саме за типом статевого процесу). У цих відділах оогамія здійснюється по різному. У хітридіомікотових нерухома яйцеклітина запліднюється рухливими сперматозоїдами, що мають джгутики, тобто представлена типовим варіантом. У оомікотових нерухомі яйцеклітини запліднюються не сперматозоїдами, а вмістом недиференційованих на клітини багатоядерних чоловічих гаметангіїв, які переливають свій вміст в яйцеклітини через спеціальні вирости клітинної оболонки.
- Гаметангіогамія являє собою процес злиття вмісту недиференційованих на гамети двох гаметангіїв. Варіанти статевого процесу цього типу відомі у зигомікотових (під назвою зигогамії) та аскомікотових. При зигогамії зливаються дві багатоядерні клітини, які розташовуються на верхівках гіф неклітинного міцелію, від якого відокремлені септами. Морфологічно ці клітини виглядають однаково, але їх ядра мають різні статеві знаки. Після плазмагамії ядра різних статевих знаків попарно зливаються, і утворюють зиготу з багатьма диплоїдними ядрами. Вона одягається багатошаровою щільною оболонкою і перетворюється на зигоспору. Після періоду спокою в зигоспорі відбувається мейоз, і вона утворює гаплоїдну гіфу, на верхівці якої формується спорангій з гаплоїдними спорангіоспорами. Гаметангіогамія з утворенням дикаріонів притаманна аскомікотовим грибам. Тут також зливаються гаметагнгії, вміст яких не диференційований на гамети. Проте, на відміну від зигогамії, плазмогамію не супроводжує каріогамія. Крім того, хоча гаметангії багатоядерні, в статевому процесі бере участь лише по одному ядру з кожного гаметангію, які утворюють дикаріон. З такої клітини злиття з дикаріонтичним ядром, як правило, розвиваються гіфи дикаріонтичного міцелію. Згодом ядра дикаріону зливаються, утворюючи диплоїдне зиготичне ядро. Далі відбувається мейоз і розвивається статеве спороношення, представлене сумкою (аском). В аску ендогенно розвиваються гаплоїдні аскоспори.
- Соматогамія, при якій також утворюються дикаріони, характерна для базидієвих грибів. Спеціалізовані статеві органи в цьому випадку повністю втрачені, а їх функцію виконують соматичні клітини вегетативного міцелію. Копуляція відбувається між двома вкритими оболонками клітинами гаплоїдних гіф. При цьому плазмагамію не супроводжує каріогамія, натомість утворюються дикаріони. З клітин з дикаріонами розвивається основний тип міцелію базидієвих — дикаріонтичний септований міцелій. Наприкінці дикаріонтичної фази ядра дикаріонів зливаються, утворюючи диплоїдне зиготичне ядро, відбувається мейоз і розвивається статеве спороношення — базидія з розташованими на її поверхні гаплоїдними базидіоспорами.
Серед грибів з соматогамним статевим процесом відомі як гомоталічні, так і гетероталічні форми. У гомоталічних грибів статевий процес може відбуватись між клітинами одного й того ж міцелію. У гетероталічних зливаються тільки клітини різних статевих знаків, які утворюються на різних міцеліях. Гетероталізм може бути двох типів: біполярний, при якому стать визначає одна пара алелів, та тетраполярний, коли стать визначають дві пари алелів, локалізовані в різних хромосомах. Такі алельні пари комбінуються незалежно, і як наслідок, клітини, які беруть участь у копуляції, мають не два (чоловіча або жіноча), а чотири статевих знаки. Таким чином, відділи грибів добре відрізняють за типом статевого процесу. Зокрема, серед слизовиків у акразіомікотових він відсутній, у міксомікотових — хологамний або ізогамний, у плазмодіофоромікотових — ізогамний. Серед псевдогрибів статевий процес у гіфохітриомікотових представлений ізогамією, а у оомікотових — оогамією, яка здійснюється без участі чоловічих гамет. У справжніх грибів в межах відділу хітридіомікотових статевий процес являє собою різні типи класичної гаметогамії — ізо-, гетеро- та оогамію; у зигомікотових — гаметангіогамію за типом зигогамії, у аскомікотових — також гаметангіогамію, але без утворення справжніх зигот. Для базидієвих притаманна соматогамія.
Статеве спороношення
Кінцевим продуктом статевого процесу у грибів є утворення диплоїдних або гаплоїдних спор, які внаслідок рекомбінації ДНК у диплоїдній фазі стають генетично відмінними від батьківських геномів. Такі спори «тиражують» результати статевого процесу. Структури, всередині яких утворюються ці спори, називають органами статевого спороношення грибів, або статевими спороношеннями. В різних відділах грибів статеві спороношення різняться, і це широко використовують в систематиці як одна з головних таксономічних ознак на рівні відділів.
Статеві спороношення відсутні у акразіомікотових слизовиків. У міксомікотових статеве спороношення представлене спорокарпами зі спорами. Спорокарпи являють собою плодові тіла, в яких утворюються спори. Статеве спороношення розвивається після статевого процесу не одразу, оскільки зигота без періоду спокою перетворюється на міксамебу або міксофлагеляту з диплоїдним ядром, і внаслідок серії мітозів або злиття з іншими амебоїдами дає початок гомо- або гетерокаріонтичному плазмодію з диплоїдними ядрами. В таких плазмодіях надалі відбувається мейоз, і плазмодії перетворюються на плодові тіла з гаплоїдними спорами.
У плазмодіофоромікотових статеве спороношення — це зооспорогенні цисти, що утворюються зі вторинного плазмодію. Подібно до міксомікотових, статеве спороношення після статевого процесу розвивається не одразу: після злиття ізогамет утворюється дикаріонтична чотириджгутикова клітина злиття, яка надалі внаслідок синхронних поділів ядер дикаріону розвивається в дикаріонтичний вторинний плазмодій. З часом ядра дикаріонів зливаються, утворюючи диплоїдне зиготичне ядро. Далі відбувається мейоз, і плазмодій повністю розпадається на одноядерні, вкриті клітинними оболонками гаплоїдні цисти, які також називають спочиваючими спорами. На відміну від міксомікотових, плазмодіофоромікотові не утворюють плодові тіла.
У оомікотових зиготи (ооспори) після періоду спокою або безпосередньо перетворюються на зооспорангій з гаплоїдними зооспорами, або проростають у спорангіофор з одним чи кількома зооспорангіями на верхівці. Таким чином, статеве спороношення оомікотових представлено зооспорангіями з зооспорами.
Статеве спороношення гіфохітриомікотових залишається остаточно не з'ясованим. Відомо, що після статевого процесу зигота збільшується у розмірах, її диплоїдне ядро багаторазово ділиться, і один з поділів є редукційним. Далі протопласт розпадається на одноядерні ділянки, подальша доля яких незрозуміла — за даними одних авторів, ці ділянки перетворюються на одноядерні спочиваючі спори, з яких урешті-решт утворюються зооспори; за даними інших дослідників, такі ділянки одразу перетворюються на зооспори. Таким чином статеве спороношення гіфохітриомікотових являє собою спорангій, але нез'ясованого типу.
Лабіринтуломікотові внаслідок статевого процесу утворюють багато диплоїдних цист. В кожній цисті відбувається мітоз, потім — мейоз, і з цисти розвивається вісім зооспор з гаплоїдними ядрами. Таким чином, статеве спороношення представлено диплоїдними зооспорогенними цистами.
У хітридіомікотових відомо два варіанти поведінки після статевого процесу. При першому варіанті (хітридіальні гриби) після статевого процесу утворюють дикаріонтичні зимові цисти; в них ядра дикаріонів при проростанні зливаються, далі редукційно діляться, після чого циста перетворюється на зооспорангій з гаплоїдними зооспорами. При другому варіанті (бластокладіальні та моноблефаридальні гриби) ядра гамет зливаються одразу, без утворення дикаріонів; зигота після періоду спокою проростає гіфою — спорофітом, на якій утворюються зооспорангії з диплоїдними зооспорами. Таким чином, статеве спороношення у хітридіомікотових представлено або зооспорогенними цистами, або зооспорангіями із зооспорами, які утворюються на гіфах спорофіту.
Статеве спороношення зигомікотових являє собою спорангій зі спорангіоспорами, який розвивається із зигоспори. Спорангіоспори нерухомі, вкриті клітинною оболонкою і мають гаплоїдний набір хромосом.
Аскомікотовим притаманне статеве спороншення, яке називається аском з аскоспорами. Після злиття гаметангіїв одна пара несестринських ядер утворює дикаріон, з якого, як правило, розвивається дикаріонтичний міцелій. В клітинах гіф такого міцелію відбувається каріогамія, і після певних морфогенетичних процесів (наприклад, з утворенням гачка, див. нижче), під час яких здійснюється редукційний поділ ядра, клітина перетворюється на ендогенний статевий спорангій — аск (сумку), всередині якої розвиваються гаплоїдні аскоспори. Аски можуть розвиватися безпосередньо на міцелії, або розташовуватись на поверхні чи всередині спеціалізованих видозмін міцелію, які називають плодовими тілами, і поділяють на закриті (клейстотеції), напіввідкриті (перитеції), відкриті (апотеції) чи несправжні (псевдотеції). Гриби, у яких плодові тіла макроскопічні, називають макроміцетами (на відміну від мікроскопічних грибів — мікроміцетів).
У базидієвих статеве спороношення представлено базидією з базидіоспорами. Після злиття цитоплазм вегетативних клітин міцелію (соматогамії), як і у аскоміцетів, каріогамія одразу не відбувається, а утворюється дикаріон, з якого розвивається дикаріонтичний міцелій. Згодом окремі клітини гіф такого міцелію починають перетворюватись на органи статевого спороношення. Під час цього процесу відбувається каріогамія, далі — мейоз, і врешті-решт з таких клітин розвивається гаплоїдна базидія. У випини, які утворює оболонка базидії, мігрують дочірні гаплоїдні ядра. Випини відділяються від базидії септами, і перетворюються на одноядерні вкриті оболонкою гаплоїдні базидіоспори. Таким чином, базидіоспори, на відміну від аскоспор, утворюються екзогенно. Базидії можуть розташовуватись безпосередньо на міцелії або на поверхні відкритих (гімнокарпних), напіввідкритих (геміангіокарпних) чи всередині закритих (ангіокарпних) плодових тіл. У багатьох базидієвих базидії розвиваються зі спочиваючих дикаріонтичних клітин — телейтоспор.
Життєві цикли
Цикломорфоз
Є єдиним типом життєвого циклу лише у акразіомікотових грибів. Здатність до статевого розмноження не виявлена або повністю втрачена у багатьох грибів, які за комплексом морфологічних, цитологічних та молекулярно-генетичних ознак належать до аскомікотових та базидіомікотових грибів. Оскільки системи цих відділів базують, в першу чергу, на ознаках, пов'язаних з органами статевого спороношення, для агамних представників не вдається визначити чіткого місця у системі цих відділів. Такі гриби об'єднані у штучну групу грибів з нез'ясованим систематичним положенням, яку сьогодні називають групою мітоспорових грибів, а також недосконалими грибами (Fungi Imperfecti) чи класом дейтероміцетів (Deuteromycetes). Мітоспорові гриби розмножуються виключно нестатевим шляхом, переважно за допомогою конідій. Таким чином, цикломорфоз є основним типом життєвого циклу у анаморфних грибів.
Хоча у більшості мітоспорових грибів статевий процес відсутній, рівень рекомбінантної мінливості є досить високим. Він обумовлений явищами гетерокаріозісу (різноядерності) та парасексуальним циклом. При гетерокаріозісі в різних клітинах одного міцелію можуть знаходитися генетично неоднакові ядра, які через анастомози переходять з однієї клітини міцелію в іншу. Хоча «мігруючі» ядра зазвичай не зливаються з ядрами клітини-реципієнта, але привносять нову генетичну інформацію і розширюють тим самим адаптивні можливості гриба по відношенню умов зовнішнього середовища.
В окремих випадках «мігруючі» ядра зливаються з ядрами клітини-реципієнта. Таке явище називають парасексуальним циклом. Воно призводить до утворення диплоїдних ядер, що забезпечують генетичні зміни грибного організму. Парасексуальний цикл включає кілька етапів: злиття ядер гетерокаріона і утворення диплоїдного гетерозиготного ядра; розмноження таких гетерозиготних ядер в міцелії; мітотична рекомбінація при розмноженні диплоїдних ядер; вегетативна гаплодізація диплоїдних ядер внаслідок втрати хромосом.
В інших відділах принаймні у окремих представників виявлено статеве розмноження, тобто існують життєві цикли зі зміною ядерних фаз. Проте конкретні типи життєвих циклів не завжди визначені через брак експериментальних даних щодо окремих стадій онтогенезу. Зокрема, така ситуація має місце для лабіринтуломікотових та гіфохітриомікотових псевдогрибів, де спостереженнями охоплені не всі стадії, незважаючи на відомості про наявність статевого процесу.
Життєві цикли евгамних грибів
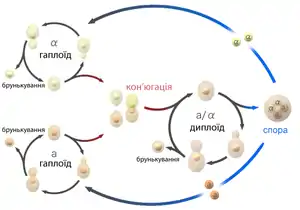
В онтогенезі евгамних грибів наявна вегетативна стадія, представлена особинами з дикаріонтичними клітинами. Після статевого процесу утворюється не зигота, а клітина злиття з дикаріонтичними ядрами. Зигота у класичному розумінні цього терміну в онтогенезі може бути взагалі відсутня, хоча замість неї після каріогамії обов'язково утворюються диплоїдні зиготичні ядра. Такі ядра є гомологами зиготи, і при розрахунку схеми життєвого циклу саме їх визначають як зиготу. Присутність у життєвому циклі грибів дикаріонтичних поколінь обов'язково відзначають у назві типу життєвого циклу. В межах конкретних відділів грибів життєві цикли за їх типом є переважно досить одноманітними, що дозволяє використовувати цю ознаку при характеристиці грибних таксонів найвищого рангу.
Для міксомікотових характерні диплофазні життєві цикли з гаметичною редукцією та без зміни поколінь або гаплодиплофазні цикли зі споричною редукцією та неправильним чергуванням ізоморфних поколінь без дикаріонтичних фаз. У плазмодіофоромікотових життєві цикли гаплофазні, з зиготичною редукцією та чергуванням одноядерного гаметофіта та дикаріонтичного спорофіта. Оомікотові гриби мають диплофазні життєві цикли з гаметичною редукцією та без зміни поколінь.
Для справжніх грибів у більшості випадків характерні життєві цикли з наявністю дикаріонтичних поколінь, і лише у частини хітридіомікотових та зигомікотових грибів такі покоління відсутні. Аскомікотові та базидіомікотові мають переважно гаплофазні життєві цикли з зиготичною редукцією та чергуванням міцеліїв з одноядерними та дикаріонтичними клітинами. У частини хітридіомікотових грибів життєвий цикл гаплофазний з зиготичною редукцією та чергуванням одноядерного плазмодія з літніми цистами та дикаріонтичного плазмодію з зимовими цистами. У іншої частини представників цього відділу життєвий цикл диплофазний з гаметичною редукцією та неправильним чергуванням поколінь і без стадій дикаріонів. Для зигомікотових характерні гаплофазні життєві цикли з зиготичною редукцією та без зміни поколінь.
Часто один і той же гриб в життєвому циклі може мати декілька типів спороношення: нестатеве та статеве, які послідовно змінюють одне одного. Таке послідовне чергування анаморф та теломорф у грибів отримало назву плеоморфізму. Явище плеоморфізму широко представлено у грибів різних відділів, проте найбільшого розвитку воно отримало у аскомікотових.
Екологія
Гриби, як і більшість інших організмів, вийшли з води (морські гриби), тому більшість водних грибів — стародавні групи: хитридіоміцети, ооміцети, гідрофохітрилієві. Вони зустрічаються у морі й прісних водоймах, утворюють рухомі спори із джутиками й живляться живими та мертвими водними організмами — водоростями, безхребетними тваринами. Однак є невелика група недосконалих грибів, які постійно живуть у прісних водоймах, предки яких жили на суші. Вони не мають зооспор й розмножуються конідіями, які мають специфічні пристосування. Конідії дуже великі, багатоклітинні, із променевими виростами або скручені спіраллю. Така форма дозволяє їм довше знаходитися у рівноважному стані у воді (подібні пристосування характерні й для планктонних водоростей) й чіплятися за субстрати (опавші у воду листя, гілки). Їх екологічна роль полягає у вивільненні зв'язаного вуглецю у прісних водоймах.
За трофічним зв'язком із субстратом гриби поділяють на еколого-трофічні групи. У найзагальнішому випадку гриби поділяють на:
Сапрофіти
Отримують поживні речовини з мертвої органічної речовини, поселяючись на різноманітних рослинних та тваринних залишках і розкладаючи їх. Гриби-паразити поселяються на інших організмах (рослинах, тваринах, інших грибах) і споживають органічну речовину хазяїна. Симбіотрофи отримують органічні речовини внаслідок симбіозу з іншими організмами (переважно вищими рослинами та водоростями), не завдаючи партнеру помітної шкоди, а навпаки, забезпечуючи його водою та мінеральними речовинами чи надаючи йому укриття.
Загально визнаними групами сапротрофів є, зокрема, гриби-ксилотрофи, які розвиваються на деревині, гумусні та підстилкові сапротрофи, що зростають у ґрунті або у лісовій підстилці, відповідно, копротрофи, що утилізують екскременти тварин тощо.
Паразитичні гриби
Серед паразитичних грибів основними групами вважають:
- фітопатогенні гриби, які паразитують на вищих рослинах — фітотрофи, альготрофи, які живляться за рахунок водоростей;
- зоотрофи, що розвиваються на різноманітних тваринах (в межах цієї групи окремо виділяють ентомофільні гриби, що уражають комах);
- мікотрофи, що паразитують на інших грибах;
- гриби-паразити людини.
За характером зв'язку з хазяїном паразитні гриби поділяють на факультативних та облігатних паразитів. Факультативні паразити для хазяїна є небезпечнішими від облігатних, оскільки спричинивши загибель господаря, продовжують свій розвиток, споживаючи його органічну речовину як сапрофіти. За топологією паразитичні гриби поділяють на внутрішньоклітинних та внутрішньотканинних паразитів.
Симбіотрофи
Найбільш відомими та численними групами грибів-симбіотрофів є мікоризоутворюючі та ліхенізовані гриби. Перші розвиваються в ризосфері вищих рослин, утворюючи зовнішню (екторофні), внутрішню (ендотрофну) або змішану мікоризи. Ліхенізовані гриби живуть у симбіозі з мікроскопічними водоростями, і утворюють симбіотичні асоціації, відомі під назвою лишайників.
Існує ряд специфічних екологічних груп грибів, які розвиваються на різноманітних матеріалах, створених людиною: металах, пластмасах, полімерних плівках, тканинах, клею, гумових виробах, склі, лакофарбових покриттях, а також на папері, книгах, рукописах, картинах тощо, зумовлюючи їх псування або біологічне пошкодження.
Культивування грибів



Гриби широко використовують як харчовий продукт, хоча їхні поживні властивості дуже малі. Але гриби цінують насамперед не за поживність, а за аромат та за специфічний смак. В культурі для задоволення потреб ринку розводять сапротрофні шапинкові гриби, такі як печериця, опеньок, глива, шіїтаке. Ці гриби не потребують особливих умов, і дають врожай плодових тіл кожні 24-48 годин, що перетворює їхнє вирощування у дуже прибуткову справу.
Культивування мікоризотворних грибів видається неможливим, оскільки для них необхідне дерево-симбіонт, інакше ріст гриба та утворення плодових тіл не відбудеться.
Для потреб лікеро-горілчаного, пивного та пекарського ринків культивують мікроскопічні пивні дріжджі, надзвичайно велику кількість штамів яких виведено методами селекції та генної інженерії. Кожен штам має окремі морфо-фізіологічні властивості, і під час бродіння виділяє у зовнішнє середовище, окрім звичайного етилового спирту та вуглекислого газу, низку специфічних речовин (цукрів, ферментів тощо), які надають кінцевій продукції особливого смаку та аромату. Наприклад, різні сорти пива отримують саме завдяки використанню різних штамів дріжджів.
Ще однією важливою цариною застосування грибів є медицина. Тут декотрі гриби використовують для отримання біологічно-активних речовин (вітамінів, антибіотиків тощо). Перший антибіотик — пеніцилін — був виділений британським вченим Александером Флемінгом з гриба Пеніциліуму зеленуватого (Penicillium viridicatum), за що йому було присуджено Нобелівську премію в галузі фізіології та медицини.
Разом із корисними для людини способами використання грибів, поширюється і зловживання потенційно отруйними та галюциногенними грибами. Зокрема, значного розповсюдження набула практика нелегального культивування сапротрофних галюциногенних грибів з роду Псилоцибе (Psilocibe). Алкалоїди псилоцибін, при інтоксикації організму, породжує сильні процеси збудження у мозку людини, які призводять до виникнення яскравих і сюрреалістичних галюцинацій.
Їстівні гриби
- Білий гриб
- Боровик жовтий
- Боровик жовто-коричневий синіючий
- Боровик королівський
- Березовик
- Веселка звичайна
- Гіропор березовий синіючий
- Гіропор каштановий
- Гігрофор золотистий
- Гігрофор модриновий
- Гігрофор пізній
- Гігрофор ранній
- Гігрофор сироїжкоподібний
- Гнойовик білий
- Гнойовик чорнильний
- Гриб зимовий
- Гриб-зонтик великий
- Гриб-зонтик червоніючий
- Дощовик їстівний
- Ентолома садова їстівна
- Заячий гриб
- Їжовик жовтуватий
- Клітоцибе бокалоподібний
- Клітоцибе величезний
- Ковпак (гриб)
- Козляк
- Лепіота червонопластинчаста
- Лепіста фіалкова
- Ліофіл зрослий
- Ліофіл скупчений
- Ліофіл травневий
- Лімацела крапчаста
- Лімацела масляна
- Лімацела ніжна
- Лисичка справжня
- Маслюк звичайний
- Маслюк зернистий
- Маслюк модриновий
- Маслюк модриновий синіючий
- Маслюк слизький
- Моховик жовто-бурий
- Моховик зелений
- Моховик різнобарвний
- Моховик тріщинуватий
- Мухомор Цезарів
- Опеньок луговий
- Опеньок осінній справжній
- Осиковик
- Павутинник каштановий слизький
- Павутинник мінливий
- Печериця звичайна
- Печериця польова
- Печериця садова
- Печериця їстівна
- Печіночниця звичайна (гриб)
- Плеврот черепичастий
- Плютей бурий
- Польський гриб
- Поліпіл зонтичний
- Порхавка гігантська
- Підберезовик
- Підвишень
- Рижик смачний
- Рядовка зелена
- Рядовка наземна
- Рядовка темно-сіра
- Рядовка тополева
- Рядовка червона
- Рядовка червонувата
- Сироїжка ароматна
- Сироїжка бездоганна
- Сироїжка болотяна
- Сироїжка біла
- Сироїжка вонюча
- Сироїжка зелена велика
- Сироїжка коричневолілова
- Сироїжка луската
- Сироїжка різнопластинчаста
- Сироїжка світло-жовта
- Сироїжка синьо-зелена
- Сироїжка чорно-пурпурова
- Сироїжка їстівна
- Спарасис кучерявий
- Трутовик сірчано-жовтий
- Хрящ-молочник перцевий
- Хрящ-молочник справжній
- Хрящ-молочник червоно-коричневий
- Часничник великий
- Часничник дрібний
- Часничник дубовий
- Limacella glioderma
Отруйні гриби
- Бліда поганка
- Мухомор
- Опеньок несправжній
- Клітоцибе восковий
- Клітоцибе знебарвлений
- Клітоцибе оранжево-червоний
- Клітоцибе червонуватий отруйний
- Іноцибе Патуйяра
- Іноцибе волокнистий
- Іноцибе звичайний
- Іноцибе коричневий
- Іноцибе піщаний
- Гігрофор конічний
- Дощовик несправжній
- Ентолома жовтувато-сиза отруйна
- Ентолома сіра отруйна
- Лепіота коричнево-червонувата
- Лепіота отруйна
- Мухомор білий смердючий
- Мухомор зелений
- Мухомор пантерний
- Мухомор пурпуровий
- Мухомор червоний
- Опеньок сірчано-жовтий несправжній
- Павутинник оранжево-червоний отруйний
- Печериця рудіюча отруйна
- Печериця темно-луската отруйна
- Рядовка тигриста отруйна
- Строчок звичайний
- Чортів гриб
- Галюциногенні гриби
Гриби в міфології
В божественне походження грибів та у їхній зв'язок з небом вірили греки, римляни, китайці, індуси, предки жителів Океанії та африканські народи. У північноамериканських індіанців ще й досі побутує повір'я, що гриби прийшли до нас із зірок. Мешканці багатьох континентів вірили в те, що гриби народжуються від грому. Давньогрецька назва одного із земляних грибів буквально означає «удар блискавки», в китайській мові є назва, яку перекладають як «гриб громового рокоту». В Монголії досі гриби називають «небесними плодами», а в Мексиці нащадки ацтеків — «божим тілом».
Зображення грибів, яких поїдали при ритуальних обрядах, знайдені на стінах храмів народів майя та у вигляді ритуальних статуеток, що належать до першого тисячоліття нашої ери.
Фантастичні зображення людиноподібних мухоморів були знайдені археологами серед наскельних зображень Чукотки.
Завдяки унікальним біосинтетичним здатностям та продуктам метаболізму грибів вони можуть мати чимале значення у майбутньому, зокрема у медицині та біотехнології.[14][15]
В країнах Східної Азії із легендарних часів знаний священний гриб безсмертя «Лінчжи», також зараз відомого як «Рейші». Опис цього гриба як цілющого засобу є у травнику міфічного давньокитайського володаря Шень-нуна, якого вважають першим «фармакологом», що передав людям зібрані та випробувані ним лікарські рослини[16][17][18].
Див. теж
Джерела
- Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts. Bot. Mar. 23: 371. 1980.
- Mueller GM, Schmit JP (2006). Fungal biodiversity: what do we know? What can we predict?. Biodivers Conserv 16: 1–5. doi:10.1007/s10531-006-9117-7.
- Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П. О. , ред. (2004). Розділ 2. Гриби. Ботаніка. Водорості та гриби (Навчальний посібник). Київ.
- David A. Poirier, Kenneth L. Feder (2001). Dangerous Places. Greenwood Publishing Group. ISBN 9780897896320. Page 71.
- Hawksworth, D.L. 1992. Fungi: a neglected component of biodiversity crucial to ecosystem function and maintenance. Canadian Biodiversity 1(4):4-10.
- Lutzoni, F., Kauff, F., Cox, C.J., McLaughlin, D., Celio, G., Dentinger, B., Padamsee, M., Hibbett, D., James, T.Y., Baloch, E., Grube, M., Reeb, V., Hofstetter, V., Schoch, C., Arnold, A.E., Miadlikowska, J., Spatafora, J., Johnson, D., Hambleton, S., Crockett, M., Shoemaker, R., Sung, G.‐H., Lücking, R., Lumbsch, T., O'Donnell, K., Binder, M., Diederich, P., Ertz, D., Gueidan, C., Hansen, K., Harris, R.C., Hosaka, K., Lim, Y.‐W., Matheny, B., Nishida, H., Pfister, D., Rogers, J., Rossman, A., Schmitt, I., Sipman, H., Stone, J., Sugiyama, J., Yahr, R. and Vilgalys, R. Assembling the fungal tree of life: progress, classification, and evolution of subcellular traits // American Journal of Botany. — 2004. — Т. 91. — С. 1446-1480. — DOI:.
- Дмитрий Леонтьев (10.04.2019). Предложена первая филогенетическая классификация миксомицетов. «Элементы»: Новости науки. Процитовано 27 жовтня 2021.(рос.)
- З.І. Мамчур. Мікологія. Альґологія // Програми ботанічних дисциплін. Частина І / Укл.: Н.О. Калінович, З. І. Мамчур, А. І. Прокопів, Л. О. Тасєнкевич.. — Львівський національний університет імені Івана Франка. — Львів, 2014. — 52 с.
- Bengtson, Stefan; Rasmussen, Birger; Ivarsson, Magnus (2017). Fungus-like mycelial fossils in 2.4-billion-year-old vesicular basalt. Nature Ecology & Evolution (1). doi:10.1038/s41559-017-0141.(англ.)
- Елена Наймарк (11.05.2017). В вулканических породах возрастом 2,4 млрд лет найдены следы древнейших грибов. Elementy.ru. Процитовано 11.05.2017. «Рассуждая о синхронности мицелиев в желваках и трещинах с окружающими их породами и помня о необходимости сначала заполнить пузырек смектитом, следует, вероятно, ориентироваться на более консервативную оценку возраста образования этих фоссилий. Так, в работе приведена оценка возраста высокотемпературного хлорита, наросшего на грибные гифы. Это 2,06 млрд лет. Чтобы оценить надежность этой оценки, выполненной по методу SHRIMP, читателю придется самостоятельно заглянуть в цитируемые статьи. Тут требуется высокий уровень профессиональной подготовки. Кроме того, нужно помнить, что это не возраст высокотемпературного хлорита, а одной из высокотемпературных стадий развития пород формации Унхелюк. Возможно ли, что образование крупнозернистого хлорита протекало на каком-то другом этапе их формирования? — да, возможно.»(рос.)
- Калинець-Мамчур З., 2011, Вступ.
- Philip Jacobs. Nematophagous fungi - an illustrated overview. Архів оригіналу за 4 березня 2016.
- Белякова Г.А. и др. - Ботаника, т.1.
- Molitoris H.-P. Mushrooms and Man in Medicine, Myth, and Religion// International Journal of Medicinal Mushrooms, Vol.3, p.97 (2001)
- Х. П. Моліторіс. Гриби в космічних дослідженнях. //Український ботанічний журнал, 1990, т.47, № 5, с. 70-77.
- Золотые персики Самарканда. Книга о чужеземных диковинах в империи Тан / Шефер Э. // М.: Наука, 1981. — 608 с. — С. 238—239, 534, 551.
- Shouzhong Yang (1998). The Divine Farmer's Materia Medica: A Translation of the Shen Nong Ben Cao Jing. Blue Poppy Press. с. 17—18. ISBN 9780936185965.
- Wachtel-Galor, Sissi; Yuen, John; Buswell, John A.; Benzie, Iris F. F. (2011). У Benzie, Iris F. F.; Wachtel-Galor, Sissi. Ganoderma lucidum (Lingzhi or Reishi): A Medicinal Mushroom. Herbal Medicine: Biomolecular and Clinical Aspects (вид. 2nd). Boca Raton (FL): CRC Press/Taylor & Francis. ISBN 978-1-4398-0713-2. PMID 22593926.
Література
- Антоняк Г. Л., Калинець-Мамчур З. І., Дудка І. О., Бабич Н. О., Панас Н. Є. Екологія грибів. — Львівський національний університет імені Івана Франка. — Львів: ЛНУ імені Івана Франка, 2013. — 628 с. — ISBN 978-617-10-0050-6.
- Зерова М. Я., Єлін Ю. Я., Коз'яков С. М. Гриби (їстівні, умовно їстівні, неїстівні, отруйні). — Видання третє, доповнене та перероблене. — К.: Урожай, 1979. — 232 с.
- Визначник грибів України: в 5 т. / Зерова М. Я. та ін. — К.: Наукова думка, 1967—1979.
- Гриби України: Атлас-довідник / За ред. В. П. Гелюти. — Київ: КМ Publishing, 2013. — 224 с.: іл. — ISBN 978-617-7489-52-7 (рецензія на видання; інтерв'ю з автором)
- Гриби України: Атлас-довідник / М. М. Сухомлин, В. В. Джаган. — 2-е вид., випр. і допов. — Київ: КМ-БУКС, 2017. — 240 с. : іл. — ISBN 617-7489-52-7.
- Гриби України / Василь Козак ; ред. В. Тарнопольський. — Тернопіль: Підручники і посібники, 2011. — 175 с. : кол. іл. — ISBN 978-966-07-1275-1
- Калинець-Мамчур З. (2011). Словник-довідник з альґології та мікології (для студ. вищ. навч. закл.). Серія «Біологічні Студії». Львів: Львівський національний університет імені Івана Франка. с. 399. ISBN 978-966-613-821-0.
- Леонтьєв Д. В., Акулов О.Ю. Загальна мікологія : Підручник для вищих навчальних закладів. — Видавнича група «Основа». — Х., 2007. — 228 с. : 375 іл. — ISBN 978-966-495-040-1.
Посилання
- М. Я. Зерова. Гриби / УРЕ
- Догадіна Т. В., Гонтова Т. М., Сербін А. Г. Гриби / Фармацевтична енциклопедія
- Світ грибів України
- Загадки про гриби
- (англ.) Time-Lapse Mushroom Video Stock Footage — уповільнена відеозйомка різноманітних процесів з життя грибів.
- Ходімо по гриби ОНЛАЙН-БІБЛІОТЕКА Товариства «Вартова башта» Пробудись! — 2005 (Watch Tower Bible and Tract Society of Pennsylvania)
- Чи не бажаєте грибочків?