Бурі водорості
Бу́рі во́дорості (Phaeophyceae Kjellman 1891) — велика група переважно морських, виключно багатоклітинних водоростей. На 2013 рік цей клас налічував близько 2000 видів, що належали приблизно до 260 родів[2].
? Бурі водоростіЧас існування: 150–0 млн р. т. пізня юра — наш час | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Fucus distichus | ||||||||||||
| Біологічна класифікація | ||||||||||||
| ||||||||||||
Порядки | ||||||||||||
|
Див. Класифікація | ||||||||||||
| Синоніми | ||||||||||||
| Fucophyceae, Melanophyceae, Phaeophyta | ||||||||||||
Посилання | ||||||||||||
| ||||||||||||
Бурі водорості в переважній більшості належать до морських біотопів, їхні зарості трапляються у літоральній та субліторальній зонах, до глибин 40—100 м. Представників класу Phaeophyceae можна побачити в усіх морях земної кулі, однак найбільша їхня концентрація — у приполярних та помірних широтах, де вони домінують на глибині від 6 до 15 м. Зазвичай бурі водорості ростуть прикріплено до скель та каміння різної величини, і лише в тихих місцях та на великій глибині вони можуть утримуватись на стулках молюсків або гравію. Серед усього розмаїття бурих водоростей представники лише 4 родів Heribaudiella, Pleurocladia, Bodanella та Sphacelaria можуть траплятися у прісних водоймах. Деякі морські види можуть потрапляти до прибережних солонуватих вод, де нерідко стають одним з основних компонентів флори солончакових боліт.
Представники групи відіграють важливу роль у морських екосистемах як одне з основних джерел органічної матерії, також їхні зарості створюють унікальне середовище існування для багатьох мешканців моря. Наприклад, представники роду Macrocystis, що можуть досягати 60 м завдовжки, формують великі підводні ліси біля узбереж Америки. Види роду Sargassum створюють навколишнє середовище тропічних вод Саргасового моря, найбільшого місця зростання бурих водоростей у тропіках. Багато видів, зокрема порядку Fucales, можна побачити в обростаннях підводних скель вздовж узбережжя.
Місце в системі живого світу
Попри те що з початку XX ст. існувало розуміння бурих водоростей як цілісної, монофілетичної групи з однорідним пігментним складом, однаковими продуктами асиміляції та схожою будовою рухомих стадій, питання їхньої філогенії залишалось невирішеним. Порівнюючи джгутиковий апарат та пігментний склад бурих водоростей із золотистими, Шерффель, а також багато інших альгологів, висловлювались за філогенетичну спорідненість цих двох таксонів[3].
Наприкінці XX століття, завдяки молекулярно-біологічним методам аналізу ДНК, було встановлено, що бурі водорості найближче споріднені з групою жовто-зелених водоростей (клас Xanthophyceae)[4]. 1998 року Bailey з колегами також ґрунтуючись на молекулярних дослідженнях ДНК, описали новий невеликий клас водоростей-страменопілів — Phaeothamniophyceae — представники якого, імовірно, і є прямими предками бурих водоростей[5].
До групи гетероконтів (синонімічна назва — страменопіли) об'єднували як організми, здатні до фотосинтезу (наприклад діатомей), так і ті, що втратили пластиди та живляться гетеротрофно (зокрема це різні групи грибоподібних організмів). Усіх їх об'єднує наявність у монадних клітинах двох морфологічно різних джгутиків. Джгутик, спрямований уперед, — пірчастий, вкритий двома рядами специфічних тричленних волосків — ретронем. Спрямований назад джгутик має гладеньку поверхню. Також спільною ознакою для всіх страменопілів є наявність трубчастих крист у мітохондріях. Окрім класу Phaeophyceae, до групи водоростей-гетероконтів належать такі класи водоростей: Chrysophyceae (золотисті), Eustigmatophyceae (евстигматові), Dictyochophyceae (силікофлагеляти), Bacillariophyceae (діатомеї), Raphidophyceae (хлоромонади), Xanthophyceae (жовто-зелені) та деякі інші.
За системою живого світу, яку 2005 року запропонували Сіна Адль (англ. Adl S.M) із колегами[6], клас бурих водоростей (Phaeophyceae Kjellman) включали до відділу страменопілів (Stramenopiles Patterson, 1989, emend. Adl et al., 2005 =Heterokontophyta van den Hoek et al., 1995). Відділ включали до царства Chromalveolata. [Note 1] Молекулярно-генетичні дослідження на початку 2000 років привели до об'єднання відділу страменопілів (=гетероконтів) разом з альвеолятами (Alveolata) та ризаріями (Rhizaria) у монофілетичну супергрупу SAR (Burki et al., 2007 =Harosa Cavalier-Smith, 2010). Наприкінці 2010-х отримала супергрупа SAR отримала ранг царства[7].
Бурі водорості є порівняно молодою групою організмів. За різними даними, час їхнього виникнення — від 150[8] до 200[9] мільйонів років тому. З погляду еволюції, бурі водорості є унікальною групою живих організмів, оскільки вони належать до тієї невеликої кількості ліній розвитку евкаріотів, яким, незалежно від інших, вдалось розвинути справжній багатоклітинний план будови тіла.
Класифікація
За систематикою 2013 року, яка ґрунтувалася на молекулярних, морфологічних, онтогенетичних критеріях, а також на основі відмінностей в ультраструктурній будові клітин, у класі Phaeophyceae виділяли 18 порядків[2]:
| Порядок | Кількість видів |
|---|---|
| Ectocarpales Bessey | 693 |
| Fucales Bory de Saint-Vincent | 524 |
| Dictyotales Bory de Saint-Vincent | 239 |
| Laminariales Migula | 130 |
| Sphacelariales Migula | 82 |
| Ralfsiales Nakamura ex Lim & Kawai | 54 |
| Desmarestiales Setchell & Gardner | 33 |
| Sporochnales Sauvageau | 29 |
| Cutleriales Bessey | 11 |
| Tilopteridales Bessey | 9 |
| Scytothamnales Peters & Clayton | 8 |
| Onslowiales Draisma & Prud'homme van Reine | 4 |
| Syringodermatales Henry | 4 |
| Asterocladales Silberfeld et al. | 3 |
| Discosporangiales Schmidt | 3 |
| Ishigeales Cho & Boo | 3 |
| Ascoseirales Petrov | 1 |
| Nemodermatales Parente et al. | 1 |
У систематиці 2021 року виділяли вже 21 порядок, які об'єднували в чотири підкласи[10]:
| Підкласи | Порядки | Кількість видів | |
|---|---|---|---|
| Fucophycidae Cavalier-Smith (1986) emend. Silberfeld et al. (2014) | 1603 | ||
| Ectocarpales Bessey | 769 | ||
| Fucales Bory | 561 | ||
| Laminariales Migula | 127 | ||
| Ralfsiales Nakamura ex P.-E.Lim & H.Kawai | 39 | ||
| Sporochnales Sauvageau | 33 | ||
| Desmarestiales Setchell & Gardner | 28 | ||
| Tilopteridales Bessey | 21 | ||
| Scytothamnales A.F.Peters & M.N.Clayton | 8 | ||
| Asterocladales T.Silberfeld, M.-F.Racault, R.L.Fletcher, A.F.Peters, F.Rousseau & B.de Reviers | 3 | ||
| Nemodermatales M.Parente, R.L.Fletcher, F.Rousseau & N.Phillips | 2 | ||
| Phaeosiphoniellales Silberfeld, F.Rousseau & Reviers | 1 | ||
| Ascoseirales Petrov | 1 | ||
| Dictyotophycidae Silberfeld, F.Rousseau & Reviers | 458 | ||
| Dictyotales Bory | 348 | ||
| Sphacelariales Migula | 100 | ||
| Syringodermatales E.C.Henry | 6 | ||
| Onslowiales Draisma & Prud'homme | 4 | ||
| Ishigeophycidae Silberfeld, F. Rousseau & Reviers | 8 | ||
| Ishigeales G.Y.Cho & S.M.Boo | 8 | ||
| Discosporangiophycidae Silberfeld, F. Rousseau & Reviers | 3 | ||
| Discosporangiales O.C.Schmidt | 3 | ||
Типи морфологічних структур тіла та будова талому
У межах класу представлено нитчастий, гетеротрихальний та справжній тканинний типи структур тіла водоростей. Усі бурі водорості, за винятком деяких видів роду Sargassum, ведуть прикріплений спосіб життя. Органами прикріплення слугують ризоїди або спеціалізовані базальні диски.
У представників із нитчастою структурою тіла таломи утворено системою однорядних розгалужених ниток. Гетеротрихальні водорості мають вигляд досить товстих шнурів. Такі шнури мають одно- або багатовісну будову. Серед бурих водоростей є представники з ефемерними, однорічними та багаторічними таломами. Значний вплив на тривалість існування талому мають екологічні фактори.

Найрізноманітніша форма талому характерна для представників із тканинним типом будови тіла. Форма талому таких водоростей може бути шкіркоподібною, кулястою, мішкоподібною, пластинчастою, кущеподібною з ребристими листкоподібними пластинками. Таломи деяких представників можуть мати спеціальні повітряні пухирці — пневматофори, що утримують «гілки» у вертикальному положенні. У найпростіших випадках талом сформований двома тканинами: корою, що утворена дрібними забарвленими клітинами з хлоропластами, і серцевиною, яка складається з великих безбарвних клітин, котрі виконують провідну та запасливу функцію.
У більш високоорганізованих представників талом утворений чотирма типами тканин: корою, меристодермою, проміжною тканиною та серцевиною. Меристодерма — це поверхнева тканина, що виконує захисну й репродуктивну функції. Її клітини зазвичай дрібні та здатні до активного поділу. У клітинах проміжної тканини накопичуються продукти асиміляції.
Типи росту
Для таломів бурих водоростей характерно 6 типів росту:
- Дифузний — більшість клітин рослинного організму здатні до поділу (напр. Ectocarpus, Petalonia).
- Верхівковий — до поділу здатні тільки клітини на верхівках талому (напр. Sphacelaria, Dictyota).
- Трихоталічний — клітини діляться, формуючи волоски над і під поверхнею талому (напр. Cutleria, Desmarestia).
- Промеристематичний — апікальна клітина не здатна до поділу, але контролює поділ меристематичних клітин, розташованих під нею (напр. Fucus).
- Інтеркалярний — у таломі наявна меристематична зона, клітини якої ділячись, наростають вгору та вниз від неї (напр. Laminaria).
- Меристодерматичний — ріст забезпечується спеціалізованою поверхневою тканиною, клітини якої діляться паралельно та перпендикулярно поверхні талому (напр. Sargassum).
Цитологічні та біохімічні особливості будови клітин
Клітинні покриви бурих водоростей утворено оболонками, які складаються з внутрішнього каркасного шару, утвореного мікрофібрилами целюлози, та зовнішнього аморфного, який містить здебільшого солі альгінової кислоти, пектинові речовини та білки. Солі альгінової кислоти — альгінати — у водних розчинах здатні до утворення гелів.
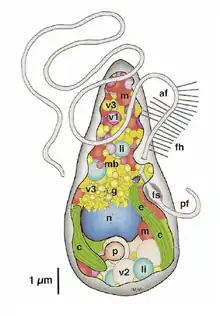
(af) передній джгутик; (c) хлоропласт; (e) вічко; (fh) ретронеми (присутні вздовж усього джгутика); (fs) базальне потовщення заднього джгутика; (g) комплекс Гольджі; (li) ліпідне включення; (m) мітохондрії; (mb) мікротільце; (mt) мікротрубочки; (n) ядро; (p) піреноїд; (pf) задній джгутик; (v1) фізоди; (v2) запасний продукт; (v3) везикули із будівельним матеріалом клітинної оболонки
Ще одним компонентом клітинних оболонок бурих водоростей є фукоїдани. За своєю хімічною природою вони є сульфатованими полісахаридами складної будови. Основний мономер фукоїданів — етерифіковані сірчаною кислотою залишки α-L-фукози. Окрім неї до складу фукоїданів можуть також входити й інші моносахариди. Подібні полісахариди не трапляються в жодній іншій групі водоростей, однак схожі на них сполуки було виявлено у морських безхребетних (морських їжаках та голотуріях)[11].
Завдяки великій кількості альгінатів та пектинових речовин, клітинні оболонки бурих водоростей здатні сильно ослизатись та перебувати в колоїдному (гелеподібному) стані.
Клітини бурих водоростей одноядерні. Ядро має типову евкаріотичну будову, усередині нього наявне велике, добре помітне ядерце. Як і в багатьох інших класів водоростей-страменопілів, з якими споріднені бурі водорості, зовнішня оболонка ядра переходить у хлоропластний ендоплазматичний ретикулюм.
Хлоропласти дрібні, численні. Мають дископодібний вигляд. У більшості бурих водоростей піреноїди в хлоропластах дуже дрібні, майже непомітні в світловий мікроскоп. Пластиди бурих водоростей вторинно-симбіотичні, родофітного типу, оточені чотирма мембранами. Дві зовнішні утворюють хлоропластний ЕПР, дві внутрішні є власне мембранами пластиди. Між зовнішніми та внутрішніми мембранами зберігається перипластидний простір. Тилакоїди у пластиді переважно розташовуються стрічками по три, на периферії розташована оперізуюча ламела.
Пігменти пластид є хлорофілами а і c, β- та ε-каротинами, а також специфічними ксантофілами — фукоксантином та віолаксантином. Саме останні й зумовлюють характерний буро-жовтий колір таломів водоростей із класу Phaeophyceae. Окрім вищезгаданих сполук, у хлоропластах бурих водоростей також присутні інші ксантофіли: діатоксантин, діадиноксантин, зеаксантин, антераксантин та неоксантин.
За межами хлоропласту, у вигляді цитоплазматичних платівок, оточених власними мембранами, відкладається основний запасний продукт бурих водоростей — специфічний полісахарид ламінарин. Окрім ламінарину, запасним продуктом у бурих водоростей може слугувати шестиатомний спирт D-маннітол. Так, у деяких видів ламінарії його концентрація восени може сягати до 25% сухої маси.
Ще однією особливістю бурих водоростей є наявність у їхніх клітинах особливих вакуолей — фізод. У світловий мікроскоп фізоди виглядають як невеликі світлозаломлюючі включення, у молодих клітинах — безбарвні, у зістарюваних — жовтого або бурого кольору. Діаметр фізод може становити від 0,1 до 10 мкм, у середньому — 1—4 мкм. На сьогодні достеменно встановлено, що вміст фізод є сукупністю поліфенольних сполук окремого класу танінів, похідних флороглюцину — флоротанінів.
Джгутикові стадії в бурих водоростей є виключно гаметами та зооспорами (у бурих водоростей відсутні рухомі вегетативні клітини). Такі клітини мають два джгутики, як і решта страменопілів. Довший джгутик — пірчастий, вкритий тричленними мастигонемами, а коротший — гладенький. Також у монадних клітин, у хлоропласті наявне вічко (стигма), що складається з 40—80 ліпідних глобул. Вічко функціонує як увігнуте дзеркало, фокусуючи світло на основі джгутика, де міститься фоторецептор, що відповідає за фототаксиси гамет або зооспор бурих водоростей.
Розмноження та життєвий цикл
У бурих водоростей спостерігаються всі типи розмноження: вегетативне, нестатеве, а також статеве. Вегетативне розмноження відбувається при випадковій фрагментації талому, і лише рід Sphacelaria формує особливі виводкові бруньки.
Нестатеве розмноження рухомими зооспорами властиве переважній більшості бурих водоростей, однак у представників порядків Dictyotales і Tilopteridales можна спостерігати розмноження нерухомими тетра- та моноспорами.
Місцем утворення зооспор слугують одно- або багатогніздні зооспорангії. Це значно збільшені у розмірах клітини, вміст яких, після серії поділів ядра, розпадається на велику кількість зооспор. Зазвичай перший поділ ядра є редукційним, тому новоутворені зооспори мають гаплоїдний набір хромосом. Потрапивши у навколишнє середовище, зооспори переходять до активного руху, але вже через кілька хвилин вони осідають на субстрат та скидають джгутики. Частина зооспор проростає в чоловічий гаметофіт, частина — в жіночий.
Статевий процес у бурих водоростей представлений ізо- та оогамією, зрідка трапляється гетерогамія.
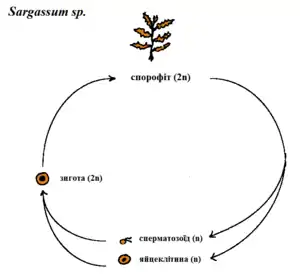
Для всього різноманіття бурих водоростей, окрім представників порядку Fucales, характерний життєвий цикл із чергуванням гаплоїдного (гаметофіт) та диплоїдного (спорофіт) поколінь. Тип життєвого циклу, в якому гаметофіт морфологічно не відрізняється від спорофіта, називають ізоморфним. У гетероморфному життєвому циклі гаметофіт бурих водоростей зазвичай має мікроскопічні розміри та виглядає як система розгалужених ниток. Продуктами гаметофіту є чоловічі та жіночі гамети, що зливаючись, дають початок новому спорофітові.
Водорості з порядку Fucales мають диплофазний життєвий цикл, без зміни поколінь. У них відсутні зооспори, мейоз відбувається при утворенні гамет, а статевий процес представлений переважно оогамією. Чоловічі та жіночі гаметангії утворюються на спеціалізованих плодючих закінченнях талому — рецептакулах. Всередині рецептакулів містяться великі напівзакриті порожнини, де розвиваються чоловічі антеридії та жіночі оогонії. Дозрілі оогонії та антеридії вивільняють яйцеклітини та сперматозоїди в навколишнє середовище, де й відбувається процес запліднення. Зигота, що утворилась, одразу ж починає ділитись та проростати в новий диплоїдний талом.
Важливу роль у статевому розмноженні бурих водоростей відіграють феромони. У бурих водоростей вони виконують дві важливі функції. По-перше — це стимуляція вивільнення чоловічих гамет, а по-друге, вони відповідальні за приваблення сперматозоїдів до жіночих гамет або яйцеклітин. За хімічною природою — це леткі, гідрофобні, лінійні або циклічні ненасичені вуглеводні. На сьогодні виявлено близько десятка феромонів у різних родів та видів бурих водоростей.
Охорона бурих водоростей
Станом на 2013 рік у Червоному Списку Міжнародного Союзу Охорони Природи (МСОП) наявні 15 видів бурих водоростей. Усі водорості, занесені до списку МСОП, ростуть біля узбережжя Галапагоських островів. Чотири з них отримали найвищу категорію вразливості «CR» (Critically Endangered). По одному виду мають категорії «EN» (Endangered) та «VU» (Vulnerable). 9 видів отримали категорію «DD» (Data Deficient), що свідчить про недостатню дослідженість динаміки популяцій цих видів. МСОП визначає кліматичні зміни (зокрема, температурний феномен Ель-Ніньйо) та пов'язане з ними надмірне розмноження організмів-фітофагів, як основні негативні фактори, що впливають на чисельність популяцій бурих водоростей[12].
До останнього видання Червоної Книги України включено 11 видів чорноморських бурих водоростей. Основною причиною скорочення їхньої чисельності є значний антропогенний вплив, що призводить до евтрофікації морських біотопів. Переважна більшість чорноморських бурих водоростей — рідкісні види з вузьколокальним або диз'юнктивним ареалом. Їхнє збереження важливе для кращого вивчення проблем біогеографії водоростей. Один вид (Sphacelaria nana) потребує охорони, оскільки є типовим представником чорноморської глибоководної флори[13].
Значення для людини
Господарське значення бурих водоростей важко переоцінити. Здавна жителі узбереж, особливо у Східній Азії, вживали їх в їжу, часто як основу раціону; сьогодні бурі водорості є передусім сировиною для отримання альгінатів, що використовуються у харчовій, текстильній, фармацевтичній та біотехнологічній галузях промисловості. У перспективі бурі водорості та їхні компоненти також розглядаються як джерело нових ліків[14].

Найбільшими країнами-виробниками продукції з бурих водоростей були й залишаються Китай, Японія та Південна Корея.
Харчова цінність
Як харчові продукти найбільшою популярністю у цих країнах (Китай, Японія та Південна Корея) користуються водорості комбу (Saccharina japonica) та вакаме (Undaria pinnatifida). Ці два види бурих водоростей здавна є об'єктами масової аквакультури в Китаї та Південній Кореї. Окрім вищезгаданих видів у Східній Азії вживаються також водорості хіджікі (Sargassum fusiforme) та араме (Eisenia bicyclis)[15]. В Україні найвідомішою харчовою бурою водорістю залишається морська капуста (Saccharina latissima).
З погляду дієтології, бурі водорості — низькокалорійна їжа з низьким вмістом жирів, збагачена вуглеводами, білками та, особливо, мінералами. Якщо порівняти зі звичайними овочами та фруктами, бурі водорості мають значно більший вміст макро- та мікроелементів, таких як натрій, калій, кальцій, магній, залізо, селен, кобальт, манган[16]. Також бурі водорості є одним з головних джерел йоду, нестача якого в організмі людини призводить до серйозних порушень розвитку, зниження працездатності, кретинізму. Останнім часом доведено, що споживання бурих водоростей та їхніх похідних є одним із найкращих методів групової та індивідуальної профілактики йододефіциту і йододефіцитних захворювань[17]. Як харчовий продукт, бурі водорості багаті на клітковину, в ролі якої виступають колоїдні компоненти їхніх клітинних оболонок — альгінати. Солі альгінової кислоти також мають значні радіопротекторні властивості[18].
Значення для науки
1907 року японський біохімік Кікуне Ікеда, досліджуючи смакові властивості традиційних страв японської кухні, приготованих із бурих водоростей комбу, описав новий тип смакових відчуттів — умамі. Із 40 кг Saccharina japonica вчений виділив 30 г глутамінової кислоти, що виявилась відповідальною за характерний смак. 1908 року компанія Ікеди розпочала промисловий випуск нової приправи — глутамату натрію[19].
2010 року міжнародною командою вчених був секвенований геном бурої водорості Ectocarpus siliculosus. Завдяки цьому, стало можливим використовувати цей організм як модельний об'єкт для вивчення питань, пов'язаних із біологією бурих водоростей, зокрема виникнення багатоклітинності[20].
Див. також
Примітки
- На час виділення царства Chromalveolata вважалося, що воно відображає окрему гілку розвитку евкаріотичних організмів, яка відокремилась на дереві життя внаслідок виняткової ендосимбіотичної події між предковою фотосинтезуючою червоною водорістю та фаготрофним евкаріотом. Із подальшою еволюцією деякі лінії живих організмів у царстві Chromalveolata втратили пластиди, тоді як інші — вторинно набули їх.
Проте за результатами пізніших досліджень об'єднання Chromalveolata було визнано штучним, немонофілетичним.
Джерела
- Kjellman, F. R. (1891). Phaeophyceae (Fucoideae). У Engler, A.; Prantl, K. Die natürlichen Pflanzenfamilien 1 (2). Leipzig: Wilhelm Engelmann. с. 176–192.
- Guiry, M.D. & Guiry, G.M. 2013. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; 02 October 2013.
- Д. К. Зеров. Очерк о филогении безсосудистых растений. — К.: Наукова думка, 1972. — 316 с.
- Ariztia, E. V.; Andersen, R. A.; Sogin, M. L. (1991). A new phylogeny of chromophyte algae using 16S-like rRNA sequences from Mallomonas papillosa (Synurophyceae) and Tribonema aequale (Xanthophyceae). Journal of Phycology 27 (3): 428–436. doi:10.1111/j.0022-3646.1991.00428.x.
- Bailey, J.C., R.R. Bidigare, S.J. Christensen and R.A. Andersen (1998). Phaeothamniophyceae classis nova.: a new lineage of chromophytes based upon photosynthetic pigments, rbcL sequence analysis and ultrastructure. Protist 149: 245–263. PMID 23194637. doi:10.1016/S1434-4610(98)70032-X.
- Adl, S. M.; et al. (2005). The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. Journal of Eukaryotic Microbiology 52 (5): 399–451. PMID 16248873. doi:10.1111/j.1550-7408.2005.00053.x.
- Остапченко, 2018, с. 59.
- Medlin, L. K.; et al. (1997). Phylogenetic relationships of the 'golden algae' (haptophytes, heterokont chromophytes) and their plastids. Plants Systematics and Evolution 11: 187–219.
- Lim, B.-L.; Kawai, H.; Hori, H.; Osawa, S. (1986). Molecular evolution of 5S ribosomal RNA from red and brown algae. Japanese Journal of Genetics 61 (2): 169–176. doi:10.1266/jjg.61.169.
- Guiry, M.D. & Guiry, G.M. Class: Phaeophyceae. AlgaeBase (Taxonomy Browser). Galway: National University of Ireland. Процитовано 29 грудня 2021.
- А. И. Усов, М. И. Билан. Фукоиданы — сульфатированые полисахариды бурых водорослей. // Успехи химии. — 2009. Том 78. — с. 846–862.
- Сайт Червоного Списку МСОП[недоступне посилання з травня 2019]
- Червона Книга України
- Albertus J. Smith. Medicinal and pharmaceutical uses of seaweed natural products: A review. // Journal of Applied Phycology. — 2004. Vol. 16. — pp. 245 −262.
- Dennis J. McHugh. A Guide to the Seaweed Industry. — Food and Agriculture Organization of the United Nations, 2003. — 106 p.
- P. Ruperez. Mineral content of edible marine seaweeds. // Food Chemistry. — 2002. No. 79. — pp. 23 — 26.
- В. Н. Корзун, А. М. Парац, А. П. Матвієнко. Проблеми і перспективи профілактики йододефіцитних захворювань у населення України. // Ендокринологія. — 2006. Т.11, № 2. — с. 187–193.
- В. Н. Корзун, В. І. Сагло, А. М. Парац. Харчування в умовах широкомасштабної аварії та її наслідків. // Український медичний часопис. — 2002. № 6 (32). — с. 99 — 105.
- Вещество с умами: вкус. — Популярная механика
- J. M. Cock et al. The Ectocarpus genome and the independent evolution of multicellularity in brown algae. // Nature — 2010.
Джерела
- Визначник прісноводних водоростей Української РСР. Вип. 12 : Червоні та бурі водорості (Rhodophyta, Phaeophyta) : у 12 т. / Н. О. Мошкова, І. О. Фролова. – К. : Наукова думка, 1983. – 208 с.
- Водоросли : справочник / Вассер С. П., Кондратьева Н. В., Масюк Н. П. и др. — К.: Наукова думка, 1989. — 608 с.
- Бойко М.Ф. Ботаніка. Систематика несудинних рослин. – Київ: Ліра-К, 2016. – C. 81-89. (PDF-файл)
- До введення до Державної Фармакопеї України монографії «Бурі водорості» / А. Г. Котов, І. М. Владимирова, В. А. Георгіянц, Л. М. Сіра // Фармаком. – 2010. – № 2. – С. 14–20. (PDF-файл)
- Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П. О. Ботаніка. Водорості та гриби: навчальний посібник. — К.: Арістей, 2006. — 476 с.
- Петров Ю.В. Отдел бурые водоросли // Жизнь растений в шести томах. Том 3. Водоросли и лишайники / Под ред. М. М. Голлербаха. — М: Просвещение, 1977. — С. 144–192.
- Robert E. Lee. Phycology. — Cambridge University Press, 2008. — 548 p.
Посилання
- Л.І. Остапченко, П.Г. Балан, Т.А. Компанець, С.Р. Рушковський. Біологія і екологія. : Рекомендовано Міністерством освіти та науки України : Підручник для 10 класу закладів загальної освіти. — Генеза. — Київ, 2018. — § 15. Огляд царств еукаріотичних організмів.
- Бурі водорості / УРЕ
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |