Кортикальна реакція
Кортикальна реакція — процес вивільнення вмісту кортикальних гранул. Відбувається під час запліднення, і є ключовим механізмом повільного блоку поліспермії у багатьох видів тварин. Сам по собі екзоцитоз кортикальних гранул, а також функціонування їх компонент мають місце і значення й поза процесом запліднення.
Механізм
Після запліднення кортикальні гранули виділяють свій вміст в перивітеліновий простір. Цей процес залежить від кальцію і опосередковується білками SNARE. Вивільнені білки модифікують позаклітинний матрикс ооцитів (наприклад, блискучу оболонку в ссавців), що в кінцевому підсумку призводить до його непроникності для сперматозоїдів.
Склад кортикальних гранул
Кількість білків кортикальних гранул ссавців залишається невідомим. За оцінками, приблизна їх кількість знаходиться в діапазоні від 4 до 14.
Вуглеводи. Кортикальні гранули ссавців містять значну кількість вуглеводів і глікозилюваних компонентів. Також вони містять α-D-ацетилгалактозамін, N-ацетилглюкозамін, N-ацетилнейрамінову кислоту, DN-ацетилгалактозамін, N-ацетилгалактозамін, та N-гліколілнейрамінову кислоту. Є свідчення, що оолемма, перивітелінового простору і блискуча оболонка модифікуються глікозильованим матеріалом, який вивільняється після запліднення і штучної активації. Ці модифікації можуть мати свій внесок в блок поліспермії або ранні стадії ембріогенезу.
Протеази. Представлені кількома ферментами. Основний внесок в блок поліспермії, як вважається, вносить модифікація компонент блискучої оболонки. В цей процес очевидно залучена трипсиноподібна протеаза, виявлена в складі кортикальних гранул. Інша серинова протеаза, активатор плазміногену тканинного типу, виявляється при заплідненні, але питання про її походження з кортикальних гранул залишається невирішеним. Також активовані овоцити виділяють протеазу, яка перетворює ZP2 в ZP2f. Як вважається, цей фермент відповідає за блок поліспермії.
Овопероксидаза. Даний фермент, судячи з ряду свідоцтв, бере участь в модифікації блискучої оболонки. Її зміцнення, ймовірно, викликається «зшиваням» залишків тирозину каталізованим даним ферментом.
Кальретикулін. Припускається, що кальретикулін служить білком-шапероном для інших компонентів кортикальних гранул, що сприяють запобіганню поліспермії. Крім того, він може сприяти запобіганню поліспермії, інгібуючи певні глікопротеїни, які сприяють взаємодії між ооцитом і спермою. З іншого боку, різні дослідження показали, що кальретікулін може виділятися з інших везикул, а не тільки з кортикальних гранул.
N- ацетилглюкозамінідаза. Являє собою глікозідазу, яка гідролізує залишки N-ацетилглюкозаміну, розташовані на блискучій оболонці. N-ацетилглюкозамін зазвичай сприяє зв'язуванню сперматозоїдів. Поки що недостатньо даних про присутність цього ферменту в овоцитах більших груп ссавців.
p32.Функціонально активний лише в ранній період запліднення або білок зазнає конформаційних змін після його вивільнення з кортикальних гранул. Ймовірно, не грає ролі в блокуванні поліспермії.
Пептидаргінін деіміназа. Присутня в кортикальних гранулах мишей і грає регулюючу роль в доімплантаційному розвитку. У блоці поліспермії, найімовірніше, також не бере участі.
Екзоцитоз кортикальних гранул
Даний процес регулюється кальцієм, а молекулярний механізм злиття мембран кортикальних гранул і плазмалеми досі залишається мало вивченим. Кортикальні гранули походженням зобов'язані апарату Гольджі і мігрують до кортексу ооцита під час оогенезу, де вони накопичуються в однорідному шарі, який зміщений на 0,4-0,6 мкм всередину від плазматичної мембрани. Білок MATER закріплює кортикальні гранули в шарі, що лежить під плазматичною мембраною яйцеклітини, де вони зв'язуються з нем'язовим міозином IIA, моторним білком. Під час запліднення і активації яйцеклітини шар кортикального актину елімінується, і кортикальні гранули переміщуються до плазматичної мембрани. Злиття кортикальних гранул з плазматичною мембраною вивільняє їх вміст, включаючи цинк і металлоендопептидазу овастацин, в позаклітинний простір. Цей вміст кортикальних гранул запобігає проникненню сперматозоїдів в матрикс оболонки і розщеплює білок zona pellucida ZP2, забезпечуючи моноспермальне запліднення[1].
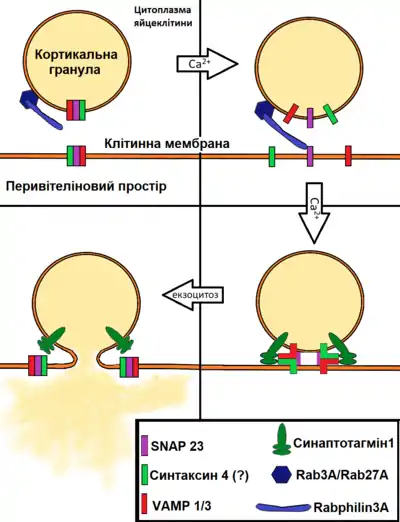
Екзоцитоз кортикальних гранул відбувається інакше, ніж інші процеси екзоцитозу в організмі, тому що кортикальні гранули не оновлюється після секреції. Вважається, що він опосередковується комплексом SNARE, який забезпечує злиття мембран і в інших екзоцитозах. Білки SNARE підрозділяються на Q (глутамін) і R (аргінін) -SNARE. Q-SNARE включають синтаксин і сімейство SNAP25, а R-SNARE включають сімейство VAMP. В ооцитах мишей синтаксин4 і SNAP23 беруть участь в екзоцитозі кортикальних гранул. Білки VAMP1 і VAMP3, як було нещодавно встановлено, беруть участь в цьому процесі[2]. З'єднання цих компонент призводить до утворення стабільного білкового комплексу, що необхідно для початку злиття мембран. За останніми даними в цьому процесі у мишей беруть участь білки α-SNAP і NSF. Також продемонстровано, що у цих тварин в кортикальній реакції бере участь білок Rab3A. Інозітол-3-фосфат є частиною кальцій-залежного сигнального шляху запуску кортикальної реакції. Він взаємодіє з ендоплазматичним ретикулумом, викликаючи вивільнення з нього депонованого кальцію. Шлях каскаду фосфатиділінозітолбіфосфата (PIP 2), опосередкованого G-білком, активація протеїнкінази C (PKC) діацілгліцеріном (DAG), також може функціонувати в регулюванні коркової реакції в ооцитах ссавців. Злиття кортикальних гранул з оолемою призводить до збільшення поверхні ооцита. Компенсаторне зменшення цієї поверхні здійснюється за рахунок індукції ендоцитозу. Кортикальна реакція вивільняє ферментативний вміст кортикальних гранул та ініціює рециркуляцию VAMP1 та дисоціацію комплексина в цитозоль.
Безхребетні
Повільний блок поліспермії у таких тварин як морські їжаки пов'язаний, як повідомляється, з активністю овопероксидази. Механізм полягає в утворенні дитирозинових містків між компонентами блискучої оболонки. Також компоненти гранул, такі як структурні білки, беруть участь в утворенні потужної оболонки навколо заплідненої яйцеклітини. Під плазмалемою ооцита морського їжака лежать близько 20 000 гранул, кожна з яких містить суміш ферментів і макромолекул. Вони вивільняють свій вміст в перивітеліновий простір після взаємодії з запліднюючим сперматозоїдом шляхом злиття з мембраною ооцита. Злиття починається в місці проникнення сперматозоїдів, а потім проходить через ооцит хвилею до антипода, що займає близько 30 секунд. Це призводить до збільшення загальної площі поверхні плазматичної мембрани, що можна спостерігати як тимчасове збільшення довжини мікроворсинок овоциту. Кортикальна реакція призводить до відокремлення мембрани запліднення і завершується під час фази реполяризації потенціалу, що виникає при заплідненні.
Значення
Кортикальна реакція еволюціонувала щоб захистити ембріон, що розвивається на ранніх стадіях ембріогенезу і забезпечити мікроклімат для ранніх стадій поділу. Також відомо, що дорсальна сторона ембріона утворюється навпроти того місця, де сперматозоїд входить в яйцеклітину. β-катенін, фактор транскрипції, і інші цитоплазматичні фактори яйцеклітини, такі як мРНК для Vg1, Xwnt11, Noggin і Activin, накопичуються на дорсальній стороні ембріона в результаті кортикальної реакції. Перші ділення зиготи стимулюються синтезом білка циклін Cdc6 з мРНК Cdc6 овоциту, що уможливлює реплікацію зиготичних хромосом за рахунок активації геліказного комплексу підтримки мініхромосом[3].
Примітки
- Vogt, EJ., Tokuhiro, K., Guo, M. et al. Anchoring cortical granules in the cortex ensures trafficking to the plasma membrane for post-fertilization exocytosis.Nat Commun 10, 2271 (2019).
- M. de Paola, F. Garrido. VAMPs sensitive to tetanus toxin are required for cortical granule exocytosis in mouse oocytes.Experimental Cell Research (2021).
- Cabej, N. R. (2019). Epigenetic Control of Early Development.Epigenetic Principles of Evolution, 119—136.
Література
- Liu M. The biology and dynamics of mammalian cortical granules. // Reproductive biology and endocrinology : RB&E. — С. 149. — DOI:.
- Hoodbhoy T, Talbot P. Mammalian cortical granules: contents, fate, and function. // Mol Reprod Dev.. — С. 439-48. — DOI:.
- Sun Q. Y. Cellular and molecular mechanisms leading to cortical reaction and polyspermy block in mammalian eggs. // Microscopy research and technique. — С. 342–348. — DOI:.
- Matilde de Paola, Oscar Daniel Bello, Marcela Alejandra Michaut. Cortical Granule Exocytosis Is Mediated by Alpha-SNAP and N-Ethilmaleimide Sensitive Factor in Mouse Oocytes // PloS one. — DOI:.
- Bello, O. D., Cappa, A. I., de Paola, M., Zanetti, M. N., Fukuda, M., Fissore, R. A., Mayorga, L. S., & Michaut, M. A. Rab3A, a possible marker of cortical granules, participates in cortical granule exocytosis in mouse eggs. // Experimental cell research. — Т. 347, вип. 1. — С. 42–51. — DOI:.
