Запліднення
Заплі́днення, стате́вий проце́с, або амфіміксис (дав.-гр. ἀμφι- — приставка зі значенням обопільності, подвійності і μῖξις — змішання) (сингамія) — процес злиття статевих клітин (гамет) рослинних і тваринних організмів, що лежить в основі статевого розмноження. Внаслідок запліднення утворюється зигота, яка дає початок новому організмові. Біологічне значення запліднення полягає у тому, що внаслідок злиття клітин з різною спадковістю при статевому розмноженні утворюється більш життєздатне потомство, ніж при нестатевому розмноженні.[1]
Статевий процес закономірно зустрічається в життєвому циклі всіх організмів, у яких відзначений мейоз. Мейоз приводить до зменшення числа хромосом вдвічі (перехід від диплоїдного стану до гаплоїдного), статевий процес — до відновлення числа хромосом (перехід від гаплоїдного стану до диплоїдного).
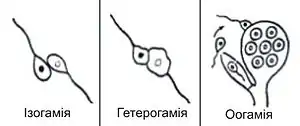
Розрізняють декілька форм статевого процесу:
- ізогамія — гамети не відрізняються одна від одної за розмірами, рухливі, джгутикові або амебоїдні;
- анізогамія (гетерогамія) — гамети відрізняються одна від одної за розмірами, але обидва типи гамет (макрогамети і мікрогамети) рухливі і мають джгутики;
- оогамія — одна з гамет (яйцеклітина) значно більша від другої, нерухома, ділення мейозу, що призводять до її утворення, різко асиметричні (замість чотирьох клітин формується одна яйцеклітина і два абортивних «полярних тільця»); інша (спермій, або сперматозоїд) рухлива, зазвичай жгутикова або амебоїдна.
Біологічне значення амфіміксиса безпосередньо пов'язано з біологічною сутністю певних сторін процесу запліднення. Дарвін, який відкрив «великий закон природи», говорив про прогресивне значення появи статевого процесу в історії органічного світу, розглядаючи при цьому перехресне запилення як джерело збагачення спадковості. Завдяки біпарієнтальній спадковості (материнське — від яйцеклітини і батьківське — від спермія) в результаті амфіміксиса виходять більш життєздатні організми, яким притаманний більш широкий спектр мінливості в порівнянні з апоміктичними рослинами.
Терміни
Іноді під статевим процесом мають на увазі не так запліднення, скільки рекомбінацію генетичної інформації між особинами одного виду, притому не обов'язково сполучену з розмноженням. У цьому випадку до різновидів статевого процесу відносять кон'югацію у протистів і парасексуальний процес у бактерій, також званий кон'югацією.
Термін амфіміксис вперше був запропонований Weismann в 1892 році. Виходячи з первісного визначення, запропонованого ним, — «змішання спадкової субстанції двох індивідуумів» цей термін підкреслює, насамперед, генетичний характер і статевий спосіб утворення нового індивідуума, тому він більш точно і більш повно відображає біологічну сутність явища ніж просто «запліднення», або «статевий процес». У багатьох підручниках і словниках[2] дається таке визначення: «амфіміксис — спосіб статевого розмноження рослин і тварин», яке є помилковим випадком вживання цього терміна як синонім до терміну «статеве розмноження».
Запліднення у тварин
Розрізняють зовнішнє запліднення, при якому злиття гамет відбувається поза тілом самки, і внутрішнє запліднення, при якому гамети зливаються у тілі самки. Зовнішнє запліднення спостерігається у багатьох безхребетних, риб, безхвостих земноводних, які відкладають статеві продукти у довкілля. Запліднення у цих тварин, а також розвиток зиготи залежить від умов середовища. У тварин із внутрішнім заплідненням (переважно наземних тварин) є спеціальні придаткові статеві органи для переносу сперми з тіла самців у тіло самок. Цей процес потребує узгодження дій самця і самки, і тому у тварин виробилися складні форми поведінки (статеві інстинкти), які забезпечують цю узгодженість.
Запліднення відбувається в результаті активного руху сперматозоїда до яйцеклітини, завдяки речовинам, що виділяються гаметами. При злитті чоловічої та жіночої статевих клітин, що мають одинарний набір хромосом, відновлюється диплоїдний набір хромосом, характерний для соматичних клітин організму. У бактерій, синьо-зелених водоростей та деяких грибів типового статевого процесу немає.
Можна виділити три фази запліднення:
- дистантна взаємодія гамет;
- контактна взаємодія гамет та активація яйцеклітини;
- злиття їх генетичного матеріалу.
Дистантна взаємодія
Дистантна взаємодія — взаємодія гамет під час осіменіння (може бути внутрішнім, якщо сперматозоїди вводяться у статеві шляхи самки, і зовнішнім, якщо гамети виводяться в зовнішнє, як правило, водне середовище). На цьому етапі організми з зовнішнім осіменінням повинні розв`язати проблему низької концентрації статевих продуктів у водному середовищі, збільшивши ймовірність зустрічі гамет, та наявності в ньому гамет різних видів, забезпечивши видоспецифічність запліднення. Тому були розвинуті механізми видоспецифічного приваблення сперматозоїдів до яйцеклітини, яке здійснюється за рахунок позитивного хемотаксису, при якому яйцеклітина секретує специфічні хемотаксичні агенти (у морського їжака були виділені сперакт та резакт), у бік більшої концентрації яких рухаються сперматозоїди.
У ссавців запліднення відбувається в ампульній частині яйцепроводу, а дистантна взаємодія здійснюється здебільшоного за рахунок реотаксису (рух сперматозоїда проти зустрічної течії рідини в маткових трубах). На цьому етапі відбувається капацитація, без якої сперматозоїд не отримає своєї запліднювальної здатності. Імовірно, це відбувається за рахунок зв`язування холестеролу і зміни холестерол/фосфоліпідного співвідношення у мембранах сперматозоїдів під дією альбуміну із секрету маткових труб. Також при цьому з галактозилтрансферази (рецептор до ZP-білків блискучої оболонки) видаляються олігоцукри, що її блокують.[3] Капацитація триває близько 7 год.[1] Сперматозоїди рухаються жіночими статевими шляхами з середньою швидкістю 2–3 мм/хв за рахунок енергії, яку вони отримують з фруктози сперми й глікозаміногліканів, які секретуються епітеліоцитами, що вистилають статеві шляхи. Ампулярної частини досягають приблизно 200 сперматозоїдів.[3]

Контактна взаємодія та активація яйцеклітини
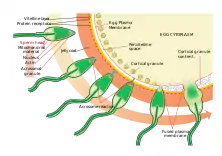
Контактна взаємодія почнається з акросомної реакції. У морського їжака вона індукується надходженням у головку спермія Са2+ та Na+ та виходом K+ і Н+, що виникають внаслідок взаємодії спермія з сульфатованими глікопротеїнами драглистої оболонки яйця; pH зростає, що підвищує активність динеїнової АТФази і , відповідно, рухливість сперматозоїда. У процесі акросомної реакції мембрана акросоми та плазмалема зливаються, вивільняючи сперміолізини. У морського їжака внаслідок полімеризації актину, що міститься між ядром і акросомою, формується акросомальна нитка (випинання задньої стінки акросоми, яка після злиття мембран опинилася на поверхні сперматозоїда). Імовірно, це відбувається за рахунок того, що підвищення pH інактивує білок, який блокував полімеризацію актину. Утворена акросомна нитка контактує з жовтковою оболонкою яйця, і біндин (видоспецифічні білки) на поверхні акросомної нитки взаємодіє з відповідним рецептором жовткової оболонки, забезпечуючи видоспецифічне впізнання гамет. Після цього яйцеклітина за допомогою полімеризації актину утворює горбик запліднення, яким "тягнеться" до акросомального виросту. Утворення горбика запліднення — це початок активації яйцеклітини. Їх мембрани зливаються і вміст сперматозоїда потрапляє до яйцеклітини. Важливу роль у злитті відіграє біндин. При цьому в мембрані яйцеклітини опиняється ділянка мембрани сперматозоїда, яка все ще зберігає високу проникність для Na+, що зумовлює деполяризацію мембрани яйцеклітини і, як наслідок, швидкий блок поліспермії. Одночасно з цим, активується фосфоліпаза С, вона розщеплює фосфатидилінозитол-3,4,5-трифосфат (мембранний фосфоліпід) з утворенням діацилгліцеролу (залишається в мембрані) та інозитолтрифосфату (міститься в цитозолі, спричиняє відкиття Са2+-каналів гладенького ендоплазматичного ретикулума). Зростання концентрації Са2+ та діацилгліцерол активують протеїнкіназу С, а та, в свою чергу, активує Na+/Н+-обмінник, який підвищує pH. Зростання pH стимулює синтез ДНК та білків. Крім того, зростання концентрації Са2+ забезпечує кортикальну реакцію (повільний блок поліспермії).[3] Кортикальний шар яйцеклітини морського їжака містить близько 15000 кортикальних гранул, кожна діаметром близько нанометра.[4] Кортикальна реакція — екзоцитоз кортикальних гранул з вивільненням сперморецепторної гідролази (фермент, що лізує сайти зв'язування сперматозоїдів та жовткової оболонки), вітелінової деламінази (фермент, що лізуює динеїнові ручки, прикріплюють жовткову оболонку до мембрани яйцеклітини), пероксидази (фермент, що "зшиває" сусідні білки жовткової оболонки, утворюючи зв`язки міє їхніми тирозиновими залишками, внаслідок оболонка твердішає і перетворюється на оболонку запліднення), глікопротеїну (зумовлює надходження рідини з яйцеклітини в утворену щілину і, як наслідок, утворення перивітелінового простору) та гіаліну (гіаліновий шар над мембраною яйцеклітини забезпечуватиме правильне розташування бластомерів при дробленні). Одночасно в яйцеклітині відбувається перерозподіл елементів цитоскелету, який має забезпечити поділи дроблення.
У ссавців акросомна нитка не утворюється. Імовірно, внаслідок спільної дії гіалуронідази і пенетрази, що вивільнилися з акросоми, та рідини яйцепроводу руйнуються контакти між фолікулярними улітинами, які утворюють променистий вінець. Биття джгутиків сперматозоїдів при цьому спричиняє обертання яйцеклітини (а точніше ооцита другого порядку, бо у ссавців, як і у більшості тварин, другий поділ мейозу завершується після контакту зі сперматозоїдом). Це обертання допомагає видалити з поверхні ооцита фолікулярні клітини. Після цього галактозилтрансфераза сперматозоїда зв`язується з N-ацетилглюкозаміном ZР3, ініціюючи акросомну реакцію. В сперматозоїд входить Са2+, він разом з Rab-білком, який теж активувався при взаємодії рецепторів сперматозоїда та ооцита, активує синаптотагміну VІ, що забезпечує злиття акросомної мембрани з плазмалемою і вивільнення ферментів. До цих ферментів належать акрозин, що формує вузький канал в прозорій оболонці, крізь який проходить сперматозоїд, та кисла фосфатаза, що забезпечує злиття мембран, якому має передувати рецепторна взаємодія.[3] У ссавців у злитті мембран беруть участь фертиліни, що містяться на мембрані сперматозоїда. Фертилін мишей зв`язується з α6β1-інтегрином на поверхні ооцита. У мишей з мунтантим фертиліном злиття не відбувається. Фертилін, як і біндин, має гідрофобний регіон, що дозволяє йому брати участь у злитті мембран.[4]
Злиття генетичного матеріалу
Залежно від виду тварини вміст сперматозоїда може весь потрапляти у яйцеклітину або лише його частина. Але мітохондрї, дистальна центріоль та джгутик сперматозоїда резорбуються у цитоплазмі яйцеклітини, тому лише незначна кількість мітохондрій в організмах походять від сперматозоїда. У мишей, за підрахунками, це лише 1 з кожних 10 000 мітохондрій. Тому можна стверджувати, що мітохондрії передаються лише від материнського організмому. Цитогамія — злиття цитоплазми яйцеклітини й сперматозоїда. У багатьох тварин одну з центріолей вносить сперматозоїд, але у мишиних зиготах обидві центріолі материнського походження. Об'єднання хромосомних наборів батьків називають каріогамією. Каріогамія може відбутися лише після завершення ооцитом поділів дозрівання, крім тих небагатьох видів, у яких сперматозоїд входить в уже зрілу яйцеклітину (морського їжак). У таких видів каріогамія відбувається шляхом безпосереднього злиття пронуклеусів і утворення єдиного ядра зиготи. У видів, ооцити яких не завершили другого мейозу до проникнення в них сперматозоїда, оболонки пронуклеусів розбираються, хромосоми розташовуються поруч в одній площині, утворюючи синкаріон, і каріогамія переходить у перший поділ зиготи. Таким чином, справжні ядра утворюються уже у двоклітинного зародка.
У морських їжаків ядро сперматозоїда потрапляє в уже зрілу яйцеклітину перпендикулярно до її поверхні. Після злиття плазматичних мембран ядро сперматозоїда та його центріоль залишаються, а мітохондрії та джгутик розпадаються всередині яйцеклітини. Потрапляючи всередину яйцеклітини, ядро сперматозоїда деконденсується, утворюючи чоловічий пронуклеус. Ядерна оболонка розпадається на невеличкі везикули, тим самим піддаючи компактний хроматин сперматозоїда дії цитоплазми яйцеклітини, внаслідок чого гістони, специфічні для сперматозоїда, заміняються на гістони, що походять з цитоплазми яйцеклітини. У морських їжаків деконденсація, імовірно, ініціюється фосфорилюванням цАМФ залежною протеїнкіназою двох специфічних для сперматозоїда гістонів. Після потрапляння сперматозоїда морського їжака потрапляє в цитоплазму яйцеклітини, чоловічий пронуклеус обертається на 180 ° так, що його центріоль розташовується між чоловічим та жіночим пронуклеусами. Потім ця центріоль діє як центр організації мікротрубочок. Ініціація синтезу ДНК відбувається на пронуклеарній стадії або після утворення ядра зиготи.
У ссавців процес пронуклеарної міграції займає близько 12 годин (у морського їжака менше години). Центріоль сперматозоїда відіграє важливу роль у заплідненні (крім зигот мишей і, можливо, деяких інших видів). Одразу ж після входження в яйцеклітину сперматозоїд повертається нею вперед у напрямку свого подальшого руху. Навколо центріолі виникає утворене мікротрубочками характерне "полярне сяйво", яке й забезпечує рух чоловічого ядра. Оболонка ядра сперматозоїда ссавців також розпадається на маленькі пухирці, що дає можливість цитоплазмі яйця "впливати" на компактний хроматин спермія: протаміни в чоловічому ядрі (наявність яких у ньому дозволяла до цього часу зберігати конденсований, неактивний стан хроматину) замінюється гістонами, хроматин деконденсується. Роль у цьому процесі відіграє руйнування дисульфідних зв’язків у протамінах глутатіоном. Хроматинова маса утримує залишки вихідної оболонки сперматозоїдного ядра на всій своїй поверхні. Невдовзі по периферії цієї маси вишикуються нові мембранні пухирці, які, з'єднуючись з ділянками попередньої оболонки, утворюючи нову оболонку чоловічого ядра, яке тепер називають чоловічим пронуклеусом. Коли сперматозод взаємодіє з ооцитом, ооцит все ще знаходиться на метафазі другого поділу мейозу (у більшості ссавців і у людини зокрема), це явище називається блокадою мейозу; для завершення мейозу необхідний сперматозоїд. Тобто звичайного ядра з ядерною оболонкою в ооциті немає, лише метафазна пластинка, що зміщена до периферії. Там завершується другий поділ мейозу і виділяється маленьке полярне тільце. Хроматин ядра яйцеклітини, після завершення поділів мейозу, також деконденсується, і жіноче ядро перетворюється на жіночий пронуклеус. Отже, після проникнення сперматозоїда в яйцеклітину, чоловічий пронуклеус ссавців набрякає, розпушується, набуває тонкої гранулярної будови, тоді як ядро ооцита завершує свій другий мейотичний поділ.
Далі пронуклеуси починають зближуватися за рахунок мікротрубочок, здійснюючи складні рухи, які називаються "танок пронуклеусів". Рух чоловічого пронуклеуса забезпечується "відштовхуванням" мікротрубочок "полярного сяйва" від поверхневого шару цитоплазми ооцита. Відрізок, який чоловічий пронуклеус проходить, рухаєючись перпендикулярно до поверхні ооцита в точці проникнення (рух не незалежить від розташування жіночого пронуклеуса) називають "доріжкою проникнення". Далі пронуклеуси починають рухаютися назустріч один одному "доріжкою копуляції". В процесі міграції пронуклеуси реплікують свою ДНК. Після зустрічі дві ядерні оболонки руйнуються. Однак замість утворення загального ядра зиготи (як це відбувається при заплідненні морського їжака), хроматин конденсується в хромосоми, які орієнтуються на загальному мітотичному веретені. Таким чином, справжнє диплоїдне ядро у ссавців спочатку спостерігається не в зиготі, а на 2-клітинній стадії.[3][5]
Запліднення в ссавців
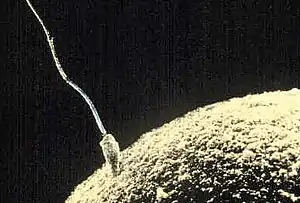
У ссавців запліднення є внутрішнім і відбувається в результаті статевого акту, коли самець вводить свій статевий орган в отвір піхви самиці. Після того, як у самця відбудеться еякуляція, велика кількість сперматозоїдів рухається до яйцеклітини.
У людей запліднення відбувається у фаллопієвих трубах за кілька годин після статевого акту. Лише один з приблизно 300 мільйонів сперматозоїдів може запліднити одну яйцеклітину. Щоб відбулося запліднення, сперматозоїд повинен увійти з піхви через шийку матки в матку і потрапити всередину однієї з фаллопієвих труб, де він має пробитись крізь щільне покриття яйцеклітини. Це покриття називається zona pellucida і складається з цукрів і білків. Кінчик головки сперматозоїда містить ензими, які проривають покриття і сприяють проникненню сперматозоїда всередину. Як тільки головка сперматозоїда опиняється всередині яйцеклітини, його «хвіст» відпадає, а її покриття додатково ущільнюється, щоб не дозволити проникненню інших сперматозоїдів.
Штучне запліднення
Ссавці, зачаті «в пробірці», почали народжуватися з 1950 років, а перша людська дитина народилась в результаті такого запліднення в 1978. З того часу — це звичайний спосіб боротьби з безпліддям.
Розрізняють два види штучного запліднення: екстраткорпоральне (поза тілом жіночої особини у загальному випадку, чи самки) та інтркорпоральне (у тілі самки).
- Екстраткорпоральне запліднення являє собою запліднення в лабораторних умовах, у тому числі так званим методом in vitro. При цьому яйцеклітини хірургічним шляхом видаляються з жіночого репродуктивного тракту і запліднюються сперматозоїдами. На другий день (4-клітинна стадія) ембріон повертають до фаллопієвої труби або матки, де його розвиток продовжується. Якщо сперматозоїди занадто слабкі, щоб проникнути до яйцеклітини, або їх вміст в спермі дуже малий, то може бути проведена операція по вживленню окремо взятого сперматозоїда прямо всередину яйцеклітини.
- Інтракорпоральне запліднення відбувається тоді, коли отримана від самця сперма використовується для вприскування у піхву або в матку самиці. В останньому випадку попередньо необхідно відділити сперматозоїди від рідини, в якій вони знаходяться. У випадку безплідності в людей також може використовуватись пересадка одразу і яйцеклітини і сперматозоїдів прямо до фаллопієвих труб, де запліднення проходить природним шляхом. Ця технологія називається англ. gamete intra-fallopian transfer (GIFT).
І яйцеклітини, і сперматозоїди можуть бути заморожені для подальшого використання під час штучного запліднення.
Запліднення у рослин
У нижчих рослин спостерігаються різні види статевого процесу: ізогамія (злиття однакових за розміром гамет), гетерогамія (злиття різних за розміром гамет), оогамія (запліднення рухливою дрібною чоловічою клітиною нерухливої і більшої за розміром жіночої), кон'югація та ін. У мохоподібних і папоротеподібних сперматозоїди, які розвиваються в антеридіях, рухаються у водному середовищі до архегонія, в якому і відбувається запліднення яйцеклітини. Вищим рослинам властива оогамія. У квіткових рослин заплідненню передує запилення. У покритонасінних відбувається так зване подвійне запліднення, що забезпечує утворення зародка та запасу поживних речовин.
У покритонасінних пилкова трубка за короткий час проходить крізь пухку тканину стовпчика і через мікропіле проникає до зародкового мішка. Ріст пилкової трубки активізує вегетативна клітина, розміщена в прикінцевій частині. За вегетативною клітиною в пилкову трубку переходить генеративна клітина, з якої в результаті мітотичного поділу утворюються два спермії. В окремих випадках спермії можуть утворюватися ще в пиляку.
В місці контакту трубки із зародковим мішком стінки останнього ослизнюються і пилкова трубка проходить усередину. Досягши яйцеклітини, пилкова трубка розривається і з неї виходять два спермії, а вегетативна клітина трубки руйнується. Спермії виходять і один із них зливається з ядром яйцеклітини, утворюючи зиготу, а другий — із вторинним (диплоїдним) ядром центральної клітини, яка дає початок ендосперму. Таким чином, у зародковому мішку відбувається відкрите професором Київського університету Святого Володимира (тепер імені Тараса Шевченка)С. Г. Навашиним (1898) подвійне запліднення. Значення подвійного запліднення полягає в тому, що і зигота, і первинна клітина ендосперму, маючи подвійну спадковість, набувають більшої життєздатності і пристосованості до умов існування. Звідси стає зрозумілою роль численних пристосувань у морфології та фізіології квітки, спрямованих на забезпечення перехресного запилення.
У зародковий мішок може проникати декілька пилкових трубок. Однак спермії цих трубок не беруть участі в заплідненні і дегенерують. Коли у зав'язі є декілька насінних зачатків, то в кожному з них відбувається описаний вище процес.
Див. також
Примітки
- Запліднення. Фармацевтична енциклопедія (укр.). Процитовано 17 березня 2021.
- Биологический словарь. — 1986. — С. 24.
- М. Е. Дзержинський, Н. В. Скрипник, О. К. Вороніна, Л. М. Пазюк (2014). Біологія індивідуального розвитку. Частина І. Практикум". Київ: "Київський університет". с. 80–88.
- Gilbert, Scott F. (2000). Gamete Fusion and the Prevention of Polyspermy. Developmental Biology. 6th edition (англ.). Процитовано 17 березня 2021.
- Gilbert, Scott F. (2000). Fusion of the Genetic Material. Developmental Biology. 6th edition (англ.). Процитовано 17 березня 2021.
Література
- Фізіологія розмноження рослин
- Біологічний словник / за ред. I. Г. Підоплічка. — К. : Головна редакція УРЕ, 1974. — Т. 3. — 552 с.
Посилання
Посилання
- Топтання // Українська мала енциклопедія : 16 кн. : у 8 т. / проф. Є. Онацький. — Накладом Адміністратури УАПЦ в Аргентині. — Буенос-Айрес, 1966. — Т. 8, кн. XV : Літери Ст — Уц. — С. 1914. — 1000 екз.
