Кінетика Міхаеліса — Ментен
Кіне́тика Міхае́ліса — Ме́нтен — у біохімії одна із найпростіших і найвідоміших моделей опису кінетики ферментативних реакцій. Говорять, що фермент слідує кінетиці Міхаеліса-Ментен, якщо для нього характерна гіперболічна залежність початкової швидкості каталізованої реакції (V0) від концентрації субстрату ([S]), що описується рівнянням (рівняння Міхаеліса — Ментен):
- ,

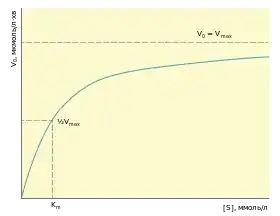
де Vmax — максимальна швидкість реакції, яка спостерігається тоді, коли фермент повністю насичений субстратом, Km — константа Міхаеліса — концентрація субстрату, при якій швидкість реакції дорівнює половині максимальної. Константа Міхаеліса-Ментен має розмірність моль/л і часто використовується для кількісного вираження спорідненості ферменту до субстрату (чим менша Km, тим більша спорідненість), проте таке її трактування не завжди коректне.
Залежність швидкості реакції від концентрацій субстрату

Одним із основних чинників, які визначають швидкість ферментативної реакції, є концентрація субстрату. Проте в експериментальних умовах вимірювання цієї залежності ускладнюється тим, що концентрація субстрату в ході реакції знижується. Тому для спрощення використовують так звану початкову швидкість V0, яка вимірюється не далі як у перші 60 с реакції, за цей час [S] встигає змінитись всього на кілька відсотків і її можна вважати наближеною до сталої величини. При незмінній концентрації ферменту і відносно невеликій концентрації субстрату швидкість реакції зростає майже лінійно при збільшенні [S] (реакція першого порядку). Проте поступово зростання стає повільнішим, аж поки при певній концентрації субстрату не виходить на плато, тобто спостерігається так званий ефект насичення. Отже після деякого значення [S] швидкість реакції наближається до максимальної Vmax і не змінюється у відповідь на подальше зростання концентрації субстрату.
Виходячи із описаних спостережень, Віктор Генрі 1903 року висловив думку про те, що зв'язування ферменту (E) із субстратом (S) із утворенням фермент-субстратного комплексу є необхідним кроком ензиматичного каталізу. Далі розвинули цю гіпотезу Леонор Міхаеліс та Мод Ментен, які 1913 року постулювали, що першим, зворотним і відносно швидким, етапом реакції є формування фермент-субстратного комплексу (ES), який на другому, повільному, етапі перетворюється у вільний фермент та продукт (P):
| . | (1) |

Оскільки друга стадія є лімітуючою, то загальна швидкість реакції повинна бути пропорційна до концентрації фермент-субстратного комплексу (речовини, що перетворюється на цій стадії).
В будь-який момент часу частина ферменту існує і вільному вигляді, тоді як інша — у зв'язаному із субстратом (ES). Співвідношення між цими двома формами залежить від [S], при малих його значеннях більшість ферменту незв'язана, зростання [S] призводить до зростання [ES] до тих пір, поки всі активні центри молекул ферменту не будуть насичені субстратом. У таких умовах буде спостерігатись максимальна швидкість реакції.
Коли фермент тільки змішується із великою кількістю субстрату, концентрація ES різко зростає, цей період називається престаціонарним і триває кілька мікросекунд, через що його дуже важко спостерігати експериментально. Після цього система досягає стаціонарного стану, при якому швидкість утворення фермент-субстратного комплексу дорівнює швидкості його утворення, тобто [ES] залишається сталою величиною. Концепція стаціонарного стану була вперше запропонована Бріґґсом та Голдейном 1925 року.
Рівняння Міхаеліса-Ментен
Виходячи із припущення, що лімітуючою стадією ферментативної реакції є розпад фермент-субстратного комплексу на вільний фермент і продукт, Міхаеліс та Ментен вивели рівняння, що описує гіперболічну криву залежності початкової швидкості від концентрації субстрату:
| (2) |

Наведене рівняння використовують тільки для односубстратних реакцій. Всі величини (V0, Vmax, Km та [S]) можна виміряти експериментально.
| Виведення рівняння Міхаеліса-Ментен | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Сучасне виведення рівняння Міхаеліса — Ментен потребує кількох припущень. По-перше, вважається, що реакція перебуває у стаціонарному стані ([ES] — стала величина). По-друге, припускається, що концентрація продукту [P] на початку реакції є мізерною, через що можна ігнорувати реакцію P + S → ES і нехтувати константою швидкості k−2. Тобто рівняння ферментативного перетворення субстрату (1) модифікується таким чином:
[E] — загальна концентрація ферменту у вільній і зв'язаній формі; Загальна швидкість реакції становить:
Проте, оскільки [ES] визначити експериментально дуже важко, потрібно виразити його через інші величини. Швидкість реакції утворення фермент-субстратного комплексу становить (нехтуючи зворотним перетворенням продукту):
Швидкість реакцій розпаду фермент-субстратного комплексу:
Для стаціонарного стану швидкість утворення фермент-субстратного комплексу рівна швидкості його розпаду, звідси:
Враховуючи що величина називається константою Міхаеліса , маємо:
Звідси концентрація фермент-субстратного комплексу становитиме:
Тепер, використовуючи рівняння (4) та (9) можна виразити швидкість реакції таким чином:
Максимальна швидкість реакції Vmax спостерігатиметься, коли весь фермент буде зв'язаний із субстратом, отже за умови, що [E] = [ES], V = Vmax. Звідси (4) перетворюється так:
Використовуючи вираз (11) можна спростити рівняння (10): |












Якщо розв'язати наведене рівняння для початкової швидкості реакції рівної половині максимальної , то отримаємо: . Отже константа Міхаеліса — це концентрація субстрату, при якій швидкість дорівнює половині максимальної.


Хоча виведення рівняння Міхеаліса — Ментен було запропоноване для простої двостадійної реакції, насправді його застосування не обмежується тільки ферментами, що діють за таким механізмом. Всі ферменти, для яких початкова швидкість реакції перебуває у гіперболічній залежності від концентрації субстрату, підкоряються кінетиці Міхаеліса — Ментен. Важливий виняток становлять регуляторні ферменти.
Хоча кінетичні константи Km та Vmax легко експериментально визначити для будь-якого ферменту, їхнє фізичне значення може суттєво відрізнятись в залежності від механізму реакції. Ці константи також не дають інформації про кількість, швидкість проходження і хімічну природу окремих кроків реакції.
Експериментальне визначення константи Міхаеліса
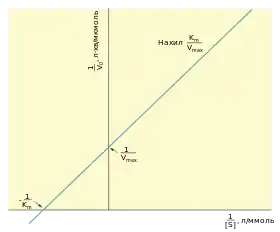
Аналіз гіперболи Міхаеліса — Ментен не дає змоги точно експериментально встановити значення Km та Vmax, вони можуть бути оцінені тільки наближено. Тому були запропоновані інші методи обчислення, найпростішим із яких є метод Лайнвівера — Берка або подвійних зворотних величин. Рівняння Лайнвівера — Берка відображає залежність 1/V0 від 1/[S]:
| ; | (12) |

Графік цього рівняння відкладається у координатах 1/V0 (вісь ординат) 1/[S] (вісь абсцис) і є лінійним. Він дозволяє легко отримати значення кінетичних констант Vmax (перетин графіка із віссю 1/V0 відповідає точці 1/Vmax) та Km (перетин графіка із віссю 1/[S] відповідає точці -1/ Km). Тангенс кута нахилу прямої становить Km/Vmax. Отримані значення будуть досить неточними, оскільки обернені графіки типу Лайнівера — Берка візуально спотворюють значення похибок вимірювань, тому в сучасних обчисленнях використовують частіше чисельні методи нелінійної регресії для прямого рівняння Міхаеліса — Ментен.
Інтерпретація кінетичних констант
Константа Міхаеліса
Значення константи Міхаеліса суттєво відрізняється для різних ферментів і навіть для різних субстратів того ж ферменту. Часто, і зазвичай помилково, його трактують як кількісну міру спорідненості ферменту до субстрату. Справжнє фізичне значення константи Міхаеліса залежить від конкретного механізму реакції (кількості етапів і відносної швидкості кожного із них). Спорідненість ферменту до субстрату характеризує константа дисоціації або субстратна константа:
| (13) |

Константа ж Міхаеліса визначається так:
| (14) |

Отже вона буде наближатись до значення субстратної константи тільки у тому випадку, коли k−1 >> k+2. Тобто Km є мірою спорідненості ферменту до субстрату тільки тоді, коли швидкість розпаду фермент-субстратного комплексу на фермент та субстрат значно переважає швидкість його розпаду на фермент і продукт. Проте в багатьох випадках k−1 << k+2 або обидві константи приблизно рівні. Навіть частіше спостерігається ситуація, коли за утворенням фермент-субстратного комплексу слідує ще серія послідовних перетворень, тоді константа Міхаеліса стає складною функцією багатьох констант швидкості.
Експериментально встановлено, що значення Km зазвичай близьке до реальної концентрації субстрату в клітині.
Максимальна швидкість і число обертів
Максимальна швидкість реакції визначається як у випадку, коли етап розпаду фермнет-субстратного комплексу на вільний фермент та продукт є лімітуючим. Проте багато кількастадійних реакцій можуть обмежуватись якимось іншим етапом. Тому зручно ввести загальну константу швидкості kcat, що описує лімітуючу стадію або стадії реакції. Наприклад для реакції (1) kcat = k+2. В загальному випадку , тоді рівняння Міхаеліса — Ментен модифікується так:

| (15) |

Константа kcat має розмірність с−1 і також називається числом обертів ферменту. Вона означає кількість молекул субстрату, що перетворюються однією молекулою ферменту за певну одиницю часу в умовах насичення ферменту субстратом.
Порівняння ефективності каталізу
Кінетичні константи Km та kcat дозволяють оцінювати ефективність каталізу певного ферменту, проте кожного з них окремо не достатньо для цього завдання. Наприклад два ферменти, що каталізують різні реакції, можуть мати однакове значення kcat, проте міра в якій вони пришвидшують відповідні реакції, може різнитись, через те, що самі реакції за відсутності ферментів можуть протікати з різною швидкістю.
Для максимально точного порівняння каталітичної ефективності різних ферментів (або одного ферменту по відношенню до різних субстратів) використовують співвідношення kcat/Km, яке ще називається константою специфічності. Це константа для перетворення E + S → E + P. Коли [S] << Km, то рівняння (15) перетворюється таким чином:
| (16) |

kcat/Km у цьому рівнянні є константою другого порядку (вимірюється у л/моль·с або М−1·c−1). Для випадку розглянутого у виведенні рівняння Міхаеліса-Ментен враховуючи визначення константи Міхаеліса із (14):
| (17) |

Отже константа специфічності буде найбільшою, коли k+2 >> k−1, тобто, коли швидкість розпаду фермнет-субстратного комплексу із утворенням продукту значно переважає над швидкістю зворотної дисоціації до вільного ферменту та субстрату. В такому випадку kcat/Km = k+1. Максимальне значення k+1 (константи другого порядку для реакції утворення фермент-субстратного комплексу) обмежене швидкістю взаємної дифузії ферменту та субстрату, тобто найбільшою кількістю разів, які вони можуть стикатись в розчині. Ця межа контрольована дифузією становить 108 М−1·c−1 — 109 М−1·c−1. Деякі ферменти, наприклад супероксиддисмутаза, каталаза, фумараза та інші, досягнули так званої каталітичної досконалості, тобто вони каталізують реакцію фактично кожен раз, коли зустрічаються із субстратом, а kcat/Km для них лежить у межах 108 М−1·c−1 — 109 М−1·c−1.
| Значення Km, kcat та kcat/Km для деяких ферментів[1][2] | ||||
|---|---|---|---|---|
| Фермент | Субстрат | Km, (моль/л) | kcat (c−1) | kcat/Km (М−1·c−1) |
| Ацетилхолінестераза | Ацетилхолін | 9,5 × 10−5 | 1,4 × 104 | 1,5 × 108 |
| Карбоангідраза | CO2 | 1,2 × 10−2 | 1,0 × 106 | 8,3 × 107 |
| HCO-3 | 2,6 × 10−2 | 4,0 × 105 | 1,5 × 107 | |
| Каталаза | H2O2 | 2,5 × 10−2 | 1,0 × 107 | 4,0 × 108 |
| Хімотрипсин | N-Ацетилгліцин етиловий естер | 4,4 × 10−1 | 5,1 × 10−2 | 1,2 × 10−1 |
| N-Ацетилвалін етиловий естер | 8,8 × 10−2 | 1,7 × 10−1 | 1,9 | |
| N-Ацетилтирозин етиловий естер | 6,6 × 10−4 | 1,9 × 102 | 2,9 × 105 | |
| Фумараза | Фумарат | 5,0 × 10−6 | 8,0 × 102 | 1,6 × 108 |
| Малат | 2,5 × 10−5 | 9,0 × 102 | 3,6 × 107 | |
| Супероксиддисмутаза | Супероксидний аніон (O2•-) | 3,6 × 10−4 | 1,0 × 106 | 2,8 × 109 |
| Уреаза | Сечовина | 2,5 × 10−2 | 1,0 × 104 | 4,0 × 105 |
| β-Лактамаза | Бензилпеніцилін | 2,0 × 10−5 | 2,0 × 103 | 1,0 × 108 |
Світлим кольором виділені ферменти із значенням kcat/Km близьким до межі, що визначається дифузією.
Примітки
- Voet et al, 2011, с. 489.
- Nelson et al, 2008, с. 199.
Джерела
- Nelson D.L., Cox M.M. (2008). Lehninger Principles of Biochemistry (вид. 5th). W. H. Freeman. с. 194—199. ISBN 978-0-7167-7108-1.
- Voet D., Voet J.G. (2011). Biochemistry (вид. 4th). Wiley. с. 487—496. ISBN 978-0470-57095-1.
- Губський Ю.І. (2007). Біологічна хімія. Київ-Вінниця: Нова книга. с. 134—137. ISBN 978-966-382-017-0.